
Корень
Историческое развитие корня
Филогенетически корень возник позже стебля и листа — в связи с переходом растений к жизни на суше и вероятно, произошёл от корнеподобных подземных веточек. У корня нет ни листьев, ни в определённом порядке расположенных почек. Для него характерен верхушечный рост в длину, боковые разветвления его возникают из внутренних тканей, точка роста покрыта корневым чехликом. Корневая система формируется на протяжении всей жизни растительного организма. Иногда корень может служить местом отложения в запас питательных веществ. В таком случае он видоизменяется.
Виды корней
Главный корень образуется из зародышевого корешка при прорастании семени. От него отходят боковые корни.
Придаточные корни развиваются на стеблях и листьях.
Боковые корни представляют собой ответвления любых корней.
Каждый корень (главный, боковые, придаточные) обладает способностью к ветвлению, что значительно увеличивает поверхность корневой системы, а это способствует лучшему укреплению растения в почве и улучшению его питания.
Типы корневых систем
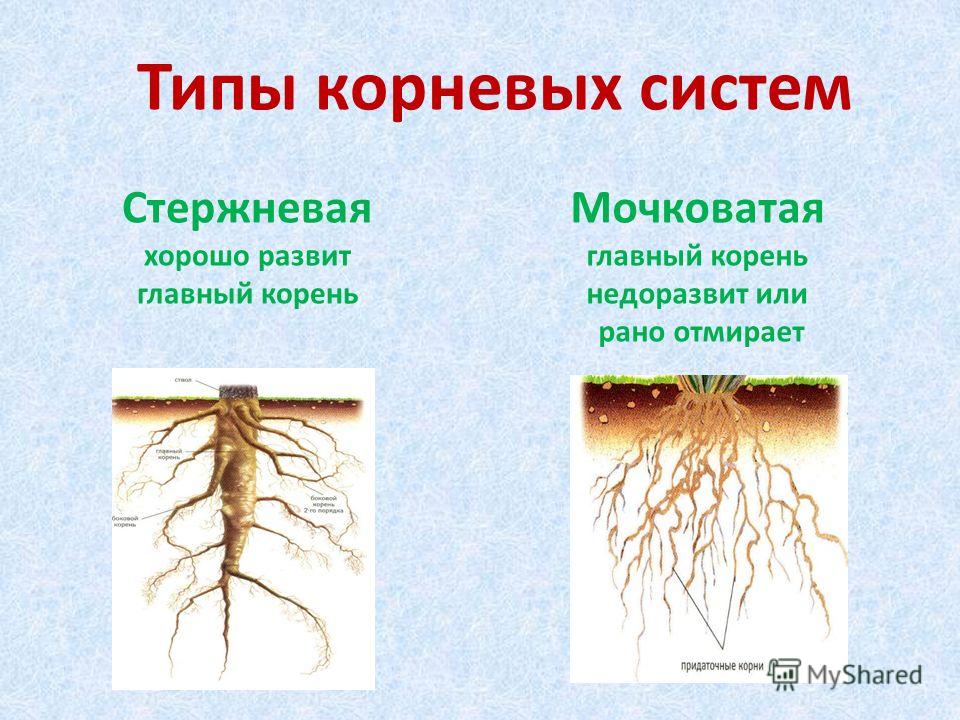
Различают два основных типа корневых систем: стержневая, имеющая хорошо развитый главный корень, и мочковатая. Мочковатая корневая система состоит из большого числа придаточных корней, одинаковых по величине. Вся масса корней состоит из боковых или придаточных корешков и имеет вид мочки.
Сильно разветвлённая корневая система образует огромную поглощающую поверхность. Например,
- общая длина корней озимой ржи достигает 600 км;
- длина корневых волосков — 10 000 км;
- общая поверхность корней — 200 м2.
Это во много раз превышает площадь надземной массы.
Если у растения хорошо выражен главный корень и развиваются придаточные корни, то формируется корневая система смешанного типа (капуста, помидор).
Внешнее строение корня. Внутреннее строение корня
Зоны корня
Корневой чехлик
Корень растёт в длину своей верхушкой, где находятся молодые клетки образовательной ткани. Растущая часть покрыта корневым чехликом, защищающим кончик корня от повреждений, и облегчает продвижение корня в почве во время роста. Последняя функция осуществляется благодаря свойству внешних стенок корневого чехлика покрываться слизью, что уменьшает трение между корнем и частичками почвы. Могут даже раздвигать частички почвы. Клетки корневого чехлика живые, часто содержат зёрна крахмала. Клетки чехлика постоянно обновляются за счёт деления. Участвует в положительных геотропических реакциях (направление роста корня к центру Земли).
Последняя функция осуществляется благодаря свойству внешних стенок корневого чехлика покрываться слизью, что уменьшает трение между корнем и частичками почвы. Могут даже раздвигать частички почвы. Клетки корневого чехлика живые, часто содержат зёрна крахмала. Клетки чехлика постоянно обновляются за счёт деления. Участвует в положительных геотропических реакциях (направление роста корня к центру Земли).
Клетки зоны деления активно делятся, протяженность этой зоны у разных видов и у разных корней одного и того же растения неодинакова.
За зоной деления расположена зона растяжения (зона роста). Протяжённость этой зоны не превышает нескольких миллиметров.
По мере завершения линейного роста наступает третий этап формирования корня — его дифференциация, образуется зона дифференциации и специализации клеток (или зона корневых волосков и всасывания). В этой зоне уже различают наружный слой эпиблемы (ризодермы) с корневыми волосками, слой первичной коры и центральный цилиндр.
Строение корневого волоска
Корневые волоски — это сильно удлинённые выросты наружных клеток, покрывающих корень. Количество корневых волосков очень велико (на 1 мм2 от 200 до 300 волосков). Их длина достигает 10 мм. Формируются волоски очень быстро (у молодых сеянцев яблони за 30-40 часов). Корневые волоски недолговечны. Они отмирают через 10-20 дней, а на молодой части корня отрастают новые. Это обеспечивает освоение корнем новых почвенных горизонтов. Корень непрерывно растёт, образуя всё новые и новые участки корневых волосков. Волоски могут не только поглощать готовые растворы веществ, но и способствовать растворению некоторых веществ почвы, а затем всасывать их. Участок корня, где корневые волоски отмерли, некоторое время способен всасывать воду, но затем покрывается пробкой и теряет эту способность.
Количество корневых волосков очень велико (на 1 мм2 от 200 до 300 волосков). Их длина достигает 10 мм. Формируются волоски очень быстро (у молодых сеянцев яблони за 30-40 часов). Корневые волоски недолговечны. Они отмирают через 10-20 дней, а на молодой части корня отрастают новые. Это обеспечивает освоение корнем новых почвенных горизонтов. Корень непрерывно растёт, образуя всё новые и новые участки корневых волосков. Волоски могут не только поглощать готовые растворы веществ, но и способствовать растворению некоторых веществ почвы, а затем всасывать их. Участок корня, где корневые волоски отмерли, некоторое время способен всасывать воду, но затем покрывается пробкой и теряет эту способность.
Оболочка волоска очень тонкая, что облегчает поглощение питательных веществ. Почти всю клетку волоска занимает вакуоль, окружённая тонким слоем цитоплазмы. Ядро находится в верхней части клетки. Вокруг клетки образуется слизистый чехол, который содействует склеиванию корневых волосков с частицами почвы, что улучшает их контакт и повышает гидрофильность системы.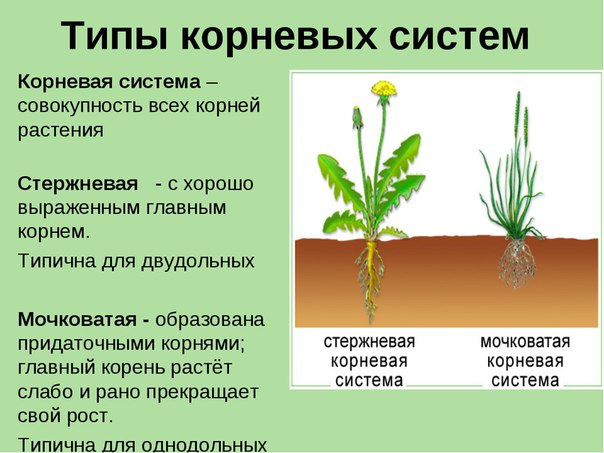 Поглощению способствует выделение корневыми волосками кислот (угольной, яблочной, лимонной), которые растворяют минеральные соли.
Поглощению способствует выделение корневыми волосками кислот (угольной, яблочной, лимонной), которые растворяют минеральные соли.
Корневые волоски играют и механическую роль — они служат опорой верхушке корня, которая проходит между частичками почвы.
Под микроскопом на поперечном срезе корня в зоне всасывания видно его строение на клеточном и тканевом уровнях. На поверхности корня — ризодерма, под ней — кора. Наружный слой коры — экзодерма, вовнутрь от неё — основная паренхима. Её тонкостенные живые клетки выполняют запасающую функцию, проводят растворы питательных веществ в радиальном направлении — от всасывающей ткани к сосудам древесины. В них же происходит синтез ряда жизненно важных для растения органических веществ. Внутренний слой коры — эндодерма. Растворы питательных веществ, поступающие из коры в центральный цилиндр через клетки эндодермы, проходят только через протопласт клеток.
Кора окружает центральный цилиндр корня. Она граничит со слоем клеток, долго сохраняющих способность к делению. Это перицикл. Клетки перицикла дают начало боковым корням, придаточным почкам и вторичным образовательным тканям. Вовнутрь от перицикла, в центре корня, находятся проводящие ткани: луб и древесина. Вместе они образуют радиальный проводящий пучок.
Это перицикл. Клетки перицикла дают начало боковым корням, придаточным почкам и вторичным образовательным тканям. Вовнутрь от перицикла, в центре корня, находятся проводящие ткани: луб и древесина. Вместе они образуют радиальный проводящий пучок.
Проводящая система корня проводит воду и минеральные вещества из корня в стебель (восходящий ток) и органические вещества из стебля в корень (нисходящий ток). Состоит она из сосудисто-волокнистых пучков. Основными слагаемыми частями пучка являются участки флоэмы (по ним вещества передвигаются к корню) и ксилемы (по которым вещества передвигаются от корня). Основные проводящие элементы флоэмы — ситовидные трубки, ксилемы — трахеи (сосуды) и трахеиды.
Процессы жизнедеятельности корня
Транспорт воды в корне
Всасывание воды корневыми волосками из почвенного питательного раствора и проведение её в радиальном направлении по клеткам первичной коры через пропускные клетки в эндодерме к ксилеме радиального проводящего пучка. Интенсивность поглощения воды корневыми волосками называется сосущей силой (S), она равна разнице между осмотическим (P) и тургорным (T) давлением: S=P-T.
Интенсивность поглощения воды корневыми волосками называется сосущей силой (S), она равна разнице между осмотическим (P) и тургорным (T) давлением: S=P-T.
Когда осмотическое давление равно тургорному (P=T), то S=0, вода перестаёт поступать в клетку корневого волоска. Если концентрация веществ почвенного питательного раствора будет выше, чем внутри клетки, то вода будет выходить из клеток и наступит плазмолиз — растения завянут. Такое явление наблюдается в условиях сухости почвы, а также при неумеренном внесении минеральных удобрений. Внутри клеток корня сосущая сила корня возрастает от ризодермы по направлению к центральному цилиндру, поэтому вода движется по градиенту концентрации (т. е. из места с большей её концентрацией в место с меньшей концентрацией) и создаёт корневое давление, которое поднимает столбик воды по сосудам ксилемы, образуя восходящий ток. Это можно обнаружить на весенних безлистных стволах, когда собирают «сок», или на срезанных пнях. Истекание воды из древесины, свежих пней, листьев, называется «плачем» растений.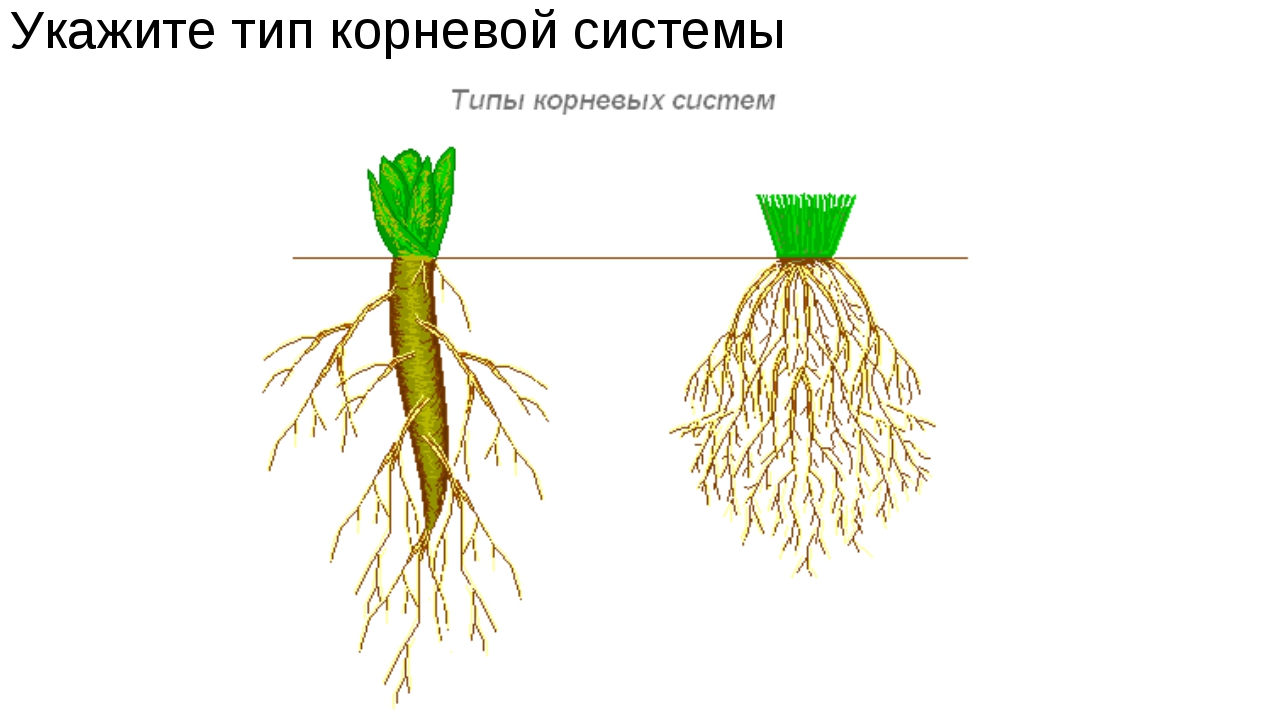 Когда распускаются листья, то они тоже создают сосущую силу и притягивают воду к себе — образуется непрерывный столбик воды в каждом сосуде — капиллярное натяжение. Корневое давление является нижним двигателем водного тока, а сосущая сила листьев — верхним. Подтвердить это можно с помощью несложных опытов.
Когда распускаются листья, то они тоже создают сосущую силу и притягивают воду к себе — образуется непрерывный столбик воды в каждом сосуде — капиллярное натяжение. Корневое давление является нижним двигателем водного тока, а сосущая сила листьев — верхним. Подтвердить это можно с помощью несложных опытов.
Всасывание воды корнями
Цель: выяснить основную функцию корня.
Что делаем: растение, выращенное на влажных опилках, отряхнём его корневую систему и опустим в стакан с водой его корни. Поверх воды для защиты её от испарения нальём тонкий слой растительного масла и отметим уровень.
Что наблюдаем: через день-два вода в ёмкости опустилась ниже отметки.
Результат: следовательно, корни всосали воду и подали её наверх к листьям.
Можно ещё проделать один опыт, доказывающий всасывание питательных веществ корнем.
Что делаем: срежем у растения стебель оставив пенёк высотой 2-3 см. На пенёк наденем резиновую трубку длиной 3 см, а на верхний конец наденем изогнутую стеклянную трубку высотой 20-25 см.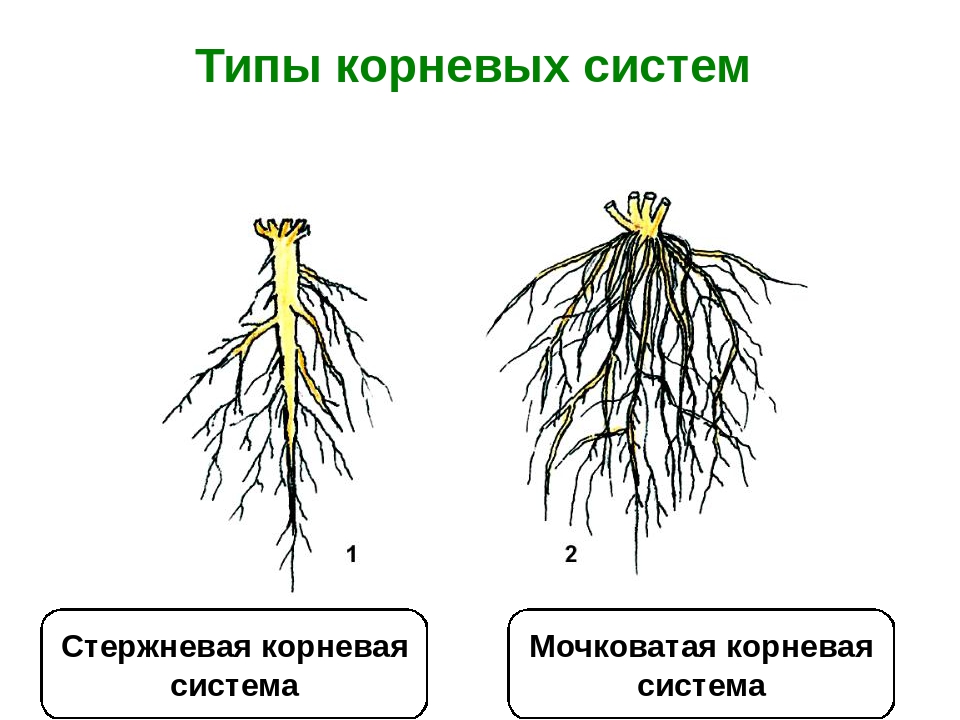
Что наблюдаем: вода в стеклянной трубке поднимается, и вытекает наружу.
Результат: это доказывает, что воду из почвы корень всасывает в стебель.
А влияет ли температура воды на интенсивность всасывания корнем воды?
Цель: выяснить, как температура влияет на работу корня.
Что делаем: один стакан должен быть с тёплой водой (+17-18ºС), а другой с холодной (+1-2ºС).
Что наблюдаем: в первом случае вода выделяется обильно, во втором — мало, или совсем приостанавливается.
Результат: это является доказательством того, что температура сильно влияет на работу корня.
Тёплая вода активно поглощается корнями. Корневое давление повышается.
Холодная вода плохо поглощается корнями. В этом случае корневое давление падает.
Минеральное питание
Физиологическая роль минеральных веществ очень велика. Они являются основой для синтеза органических соединений, а также факторами, которые изменяют физическое состояние коллоидов, т. е. непосредственно влияют на обмен веществ и строение протопласта; выполняют функцию катализаторов биохимических реакций; воздействуют на тургор клетки и проницаемость протоплазмы; являются центрами электрических и радиоактивных явлений в растительных организмах.
е. непосредственно влияют на обмен веществ и строение протопласта; выполняют функцию катализаторов биохимических реакций; воздействуют на тургор клетки и проницаемость протоплазмы; являются центрами электрических и радиоактивных явлений в растительных организмах.
Установлено, что нормальное развитие растений возможно только при наличии в питательном растворе трёх неметаллов — азота, фосфора и серы и — и четырёх металлов — калия, магния, кальция и железа. Каждый из этих элементов имеет индивидуальное значение и не может быть заменён другим. Это макроэлементы, их концентрация в растении составляет 10-2–10%. Для нормального развития растений нужны микроэлементы, концентрация которых в клетке составляет 10-5–10-3%. Это бор, кобальт, медь, цинк, марганец, молибден др. Все эти элементы есть в почве, но иногда в недостаточном количестве. Поэтому в почву вносят минеральные и органические удобрения.
Растение нормально растёт и развивается в том случае, если в окружающей корни среде будут содержаться все необходимые питательные вещества. Такой средой для большинства растений является почва.
Такой средой для большинства растений является почва.
Дыхание корней
Для нормального роста и развития растения необходимо чтобы к корню поступал свежий воздух. Проверим, так ли это?
Цель: нужен ли воздух корню?
Что делаем: возьмём два одинаковых сосуда с водой. В каждый сосуд поместим развивающие проростки. Воду в одном из сосудов каждый день насыщаем воздухом с помощью пульверизатора. На поверхность воды во втором сосуде нальём тонкий слой растительного масла, так как оно задерживает поступление воздуха в воду.
Что наблюдаем: через некоторое время растение во втором сосуде перестанет расти, зачахнет, и в конце концов погибнет.
Результат: гибель растения наступает из-за недостатка воздуха, необходимого для дыхания корня.
Видоизменения корней
У некоторых растений в корнях откладываются запасные питательные вещества. В них накапливаются углеводы, минеральные соли, витамины и другие вещества. Такие корни сильно разрастаются в толщину и приобретают необычный внешний вид. В формировании корнеплодов участвуют и корень, и стебель.
Корнеплоды
Если запасные вещества накапливаются в главном корне и в основании стебля главного побега, образуются корнеплоды (морковь). Растения, образующие корнеплоды, в основном двулетники. В первый год жизни они не цветут и накапливают в корнеплодах много питательных веществ. На второй — они быстро зацветают, используя накопленные питательные вещества и образуют плоды и семена.
Корневые клубни
У георгина запасные вещества накапливаются в придаточных корнях, образуя корневые клубни.
Бактериальные клубеньки
Своеобразно изменены боковые корни у клевера, люпина, люцерны. В молодых боковых корешках поселяются бактерии, что способствует усвоению газообразного азота почвенного воздуха. Такие корни приобретают вид клубеньков. Благодаря этим бактериям эти растения способны жить на бедных азотом почвах и делать их более плодородными.
Ходульные
У пандуса, произрастающего в приливно-отливной зоне, развиваются ходульные корни. Они высоко над водой удерживают на зыбком илистом грунте крупные облиственные побеги.
Они высоко над водой удерживают на зыбком илистом грунте крупные облиственные побеги.
Воздушные
У тропических растений, живущих на ветвях деревьев, развиваются воздушные корни. Они часто встречаются у орхидей, бромелиевых, у некоторых папоротников. Воздушные корни свободно висят в воздухе, не достигая земли и поглощая попадающую на них влагу от дождя или росы.
Втягивающие
У луковичных и клубнелуковичных растений, например у крокусов, среди многочисленных нитевидных корней имеется несколько более толстых, так называемых втягивающих, корней. Сокращаясь, такие корни втягивают клубнелуковицу глубже в почву.
Столбовидные
У фикуса развиваются столбовидные надземные корни, или корни-подпорки.
Почва как среда обитания корней
Почва для растений является средой, из которой оно получает воду и элементы питания. Количество минеральных веществ в почве зависит от специфических особенностей материнской горной породы, деятельности организмов, от жизнедеятельности самих растений, от типа почвы.
Почвенные частицы конкурируют с корнями за влагу, удерживая её своей поверхностью. Это так называемая связанная вода, которая подразделяется на гигроскопическую и плёночную. Удерживается она силами молекулярного притяжения. Доступная растению влага представлена капиллярной водой, которая сосредоточена в мелких порах почвы.
Между влагой и воздушной фазой почвы складываются антагонистические отношения. Чем больше в почве крупных пор, тем лучше газовый режим этих почв, тем меньше влаги удерживает почва. Наиболее благоприятный водно-воздушный режим поддерживается в структурных почвах, где вода и воздух находятся одновременно и не мешают друг другу — вода заполняет капилляры внутри структурных агрегатов, а воздух — крупные поры между ними.
Характер взаимодействия растения и почвы в значительной степени связан с поглотительной способностью почвы — способностью удерживать или связывать химические соединения.
Микрофлора почвы разлагает органические вещества до более простых соединений, участвует в формировании структуры почвы. Характер этих процессов зависит от типа почвы, химического состава растительных остатков, физиологических свойств микроорганизмов и других факторов. В формировании структуры почвы принимают участие почвенные животные: кольчатые черви, личинки насекомых и др.
Характер этих процессов зависит от типа почвы, химического состава растительных остатков, физиологических свойств микроорганизмов и других факторов. В формировании структуры почвы принимают участие почвенные животные: кольчатые черви, личинки насекомых и др.
В результате совокупности биологических и химических процессов в почве образуется сложный комплекс органических веществ, который объединяют термином «гумус».
Метод водных культур
В каких солях нуждается растение, и какое влияние оказывают они на рост и развитие его, было установлено на опыте с водными культурами. Метод водных культур — это выращивание растений не в почве, а в водном растворе минеральных солей. В зависимости от поставленной цели в опыте можно исключить отдельную соль из раствора, уменьшить или увеличить ее содержание. Было выяснено, что удобрения, содержащие азот, способствуют росту растений, содержащие фосфор — скорейшему созреванию плодов, а содержащие калий — быстрейшему оттоку органических веществ от листьев к корням.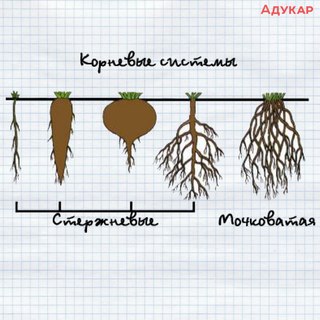 В связи с этим содержащие азот удобрения рекомендуется вносить перед посевом или в первой половине лета, содержащие фосфор и калий — во второй половине лета.
В связи с этим содержащие азот удобрения рекомендуется вносить перед посевом или в первой половине лета, содержащие фосфор и калий — во второй половине лета.
С помощью метода водных культур удалось установить не только потребность растения в макроэлементах, но и выяснить роль различных микроэлементов.
В настоящее время известны случаи, когда выращивают растения методами гидропоники и аэропоники.
Гидропоника — выращивание растений в сосудах, заполненных гравием. Питательный раствор, содержащий необходимые элементы, подаётся в сосуды снизу.
Аэропоника — это воздушная культура растений. При этом способе корневая система находится в воздухе и автоматически (несколько раз в течение часа) опрыскивается слабым раствором питательных солей.
* * *
Типы корневых систем
☰
Корневой системой называют все корни растения. Ее формируют главный корень, боковые корни и придаточные корни. Главный корень растения развивается из зародышевого корешка.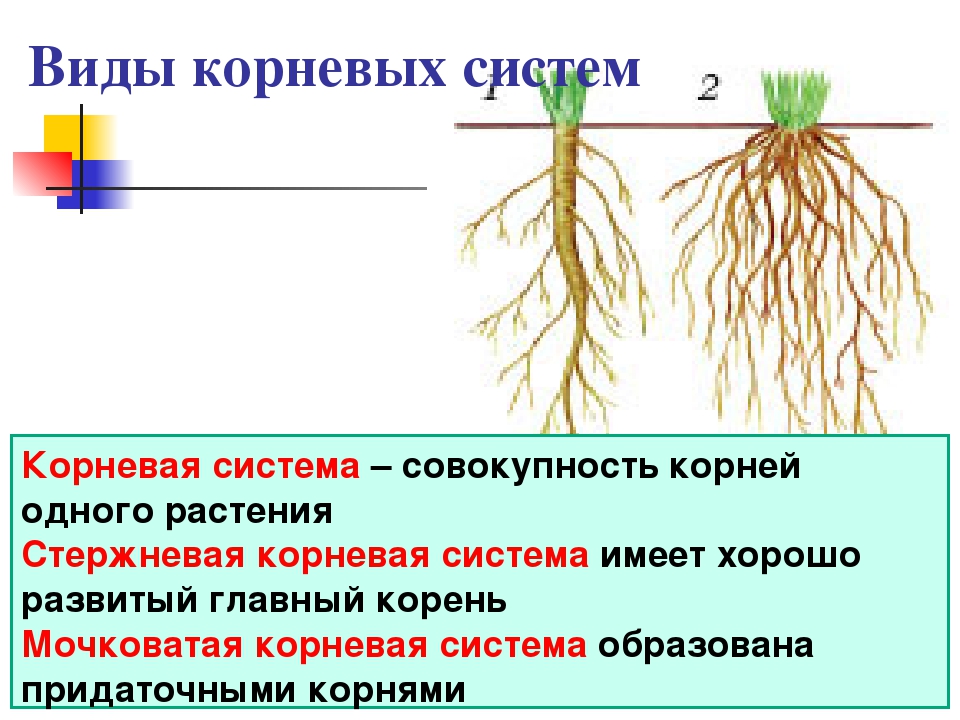 Придаточные корни обычно отрастают от нижних частей стебля растения. Боковые корни развиваются на главном и придаточном корнях.
Придаточные корни обычно отрастают от нижних частей стебля растения. Боковые корни развиваются на главном и придаточном корнях.
Корневая система растений выполняет две основные функции. Во-первых, она удерживает растение в почве. Во вторых, корни всасывают из почвы необходимые растению воду и растворенные в ней минеральные вещества.
Если у растения развивается мощный главный корень, то формируется стержневая корневая система. Если же главный корень остается неразвитым или отмирает, а развитие получают придаточные корни, то у растения формируется мочковатая корневая система.
Стержневой тип корневой системы
Стержневую корневую систему формирует не только главный корень, но и отходящие от него маленькие боковые корни.
Стержневая корневая система характерна для многих двудольных растений. Хорошо развитый главный корень есть у фасоли, клевера, подсолнечника, моркови, одуванчика.
Однако у многих многолетних растений с изначальной стержневой корневой системой рано или поздно главный корень отмирает. Вместо него от стебля отрастают многочисленные придаточные корни.
Существует подтип стержневой корневой системы — ветвистая корневая система. В этом случае сильное развитие получают несколько боковых корней. В то время как главный корень остается укороченным. Тип ветвистой корневой системы характерен для многих деревьев. Такая корневая система позволяет прочно удерживать мощные ствол и крону дерева.
Стержневая корневая система проникает в почву глубже, чем мочковатая.
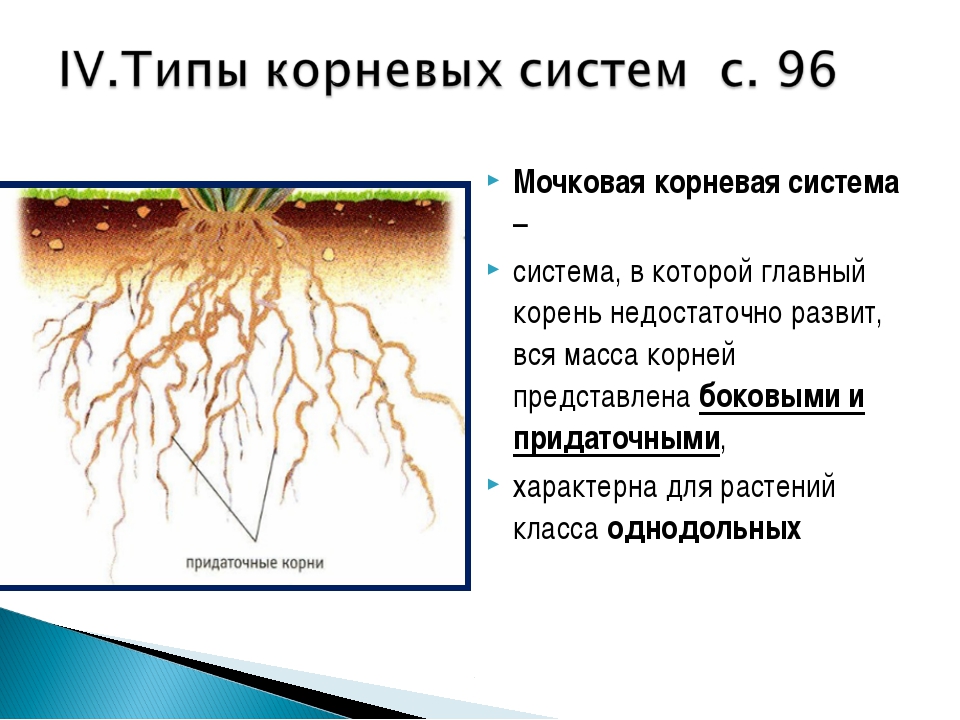
Мочковатый тип корневой системы
Для мочковатой корневой системы характерно наличие множества приблизительно одинаковых придаточных корней, которые формируют своеобразный пучок.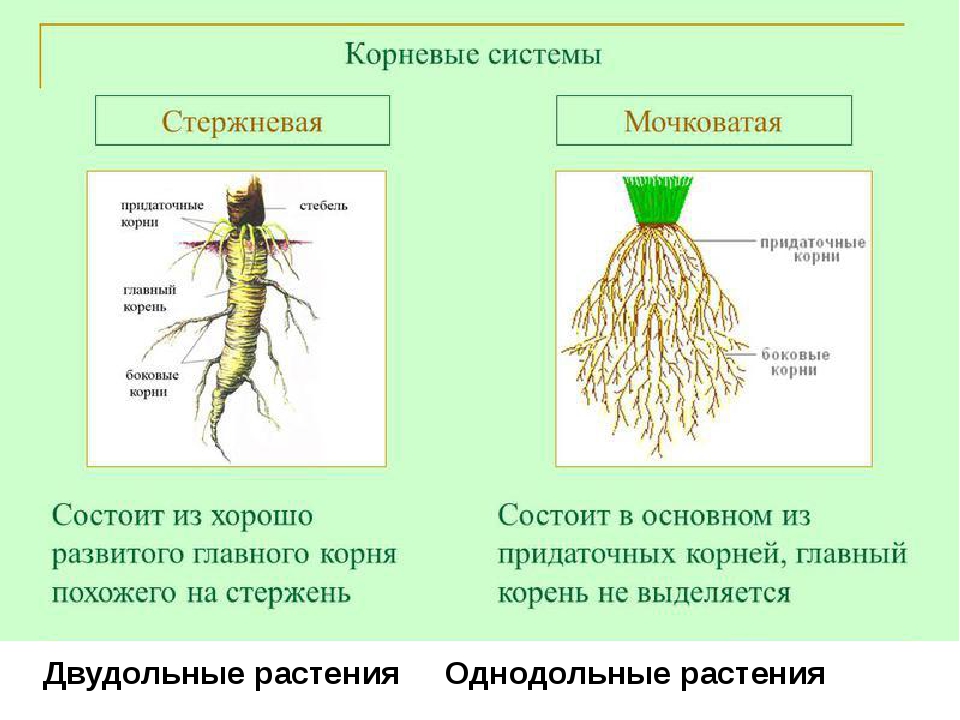 Придаточные корни отрастают от надземных и подземных частей стебля, реже от листьев.
Придаточные корни отрастают от надземных и подземных частей стебля, реже от листьев.
У растений с мочковатой корневой системой может быть и живой главный корень. Однако если он сохраняется, то не отличается по размеру от остальных корней.
Мочковатая корневая система характерна для многих однодольных растений. Среди них пшеница, рож, лук, чеснок, кукуруза, картофель.
Мочковатая корневая система хотя не проникает в почву так глубоко как стержневая, но она занимает большую площадь у поверхности почвы и плотнее оплетает частицы грунта, что улучшает всасывание водного раствора.
Видоизменения корней
☰
У корней растений есть две основные функции: 1) закрепление растения в почве, 2) поглощение воды и растворенных в ней минеральных веществ. Однако нередко корни растений выполняют ряд дополнительных функций, в результате чего видоизменяются, т. е. в природе имеют место видоизменения корней.
Ниже перечислены наиболее известные видоизменения корней.
Корнеплоды
Корнеплод — это видоизменение главного корня и нижней части стебля, в которых накапливаются запасные питательные вещества (крахмал, сахара и др.). Корнеплоды характерны для таких растений как свекла и морковь, а также ряда других.
Чаще всего корнеплоды встречаются у двулетних растений. Они образуются в конце первого вегетационного периода такого растения, обычно в конце лета или начале осени. Растение за лето накапливает запасные питательные вещества, осенью ее надземная часть отмирает. На второй год весной стебли и листья снова отрастают. При этом используются запасные вещества из корнеплода. В этот год растение цветет и плодоносит, после чего отмирает полностью.
Корнеплоды многих растений человек использует для своего питания. Урожай собирают в первый год. Если надо получить семена, то корнеплод оставляют в почве на второй год.
Корневые клубни
Корневые клубни — это видоизменения придаточных и боковых корней, в которых накапливаются запасные питательные вещества.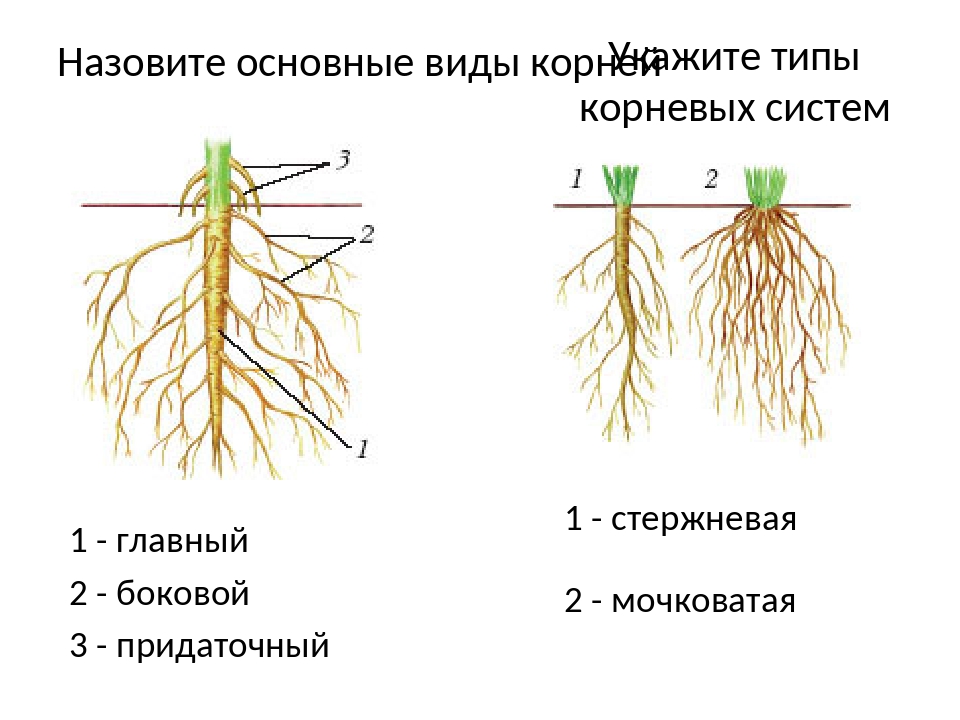 Клубни характерны для таких растений как батат, георгины и ряд других.
Клубни характерны для таких растений как батат, георгины и ряд других.
Корневые клубни по-другому называют корневыми шишками.
Корни-прицепки
Корни-прицепки (или корни-зацепки) — это видоизменения придаточных корней, служащие растению для прикрепления к какой-либо опоре. Эти корни находятся над почвой. Так растение выносит свои вегетативные части (стебель и листья) к свету при отсутствии прочного стебля. Корни-прицепки можно наблюдать у плюща.
Корни-подпорки
Корни-подпорки, или опорные корни, также развиваются из придаточных корней и находятся в воздушной среде. Они образуются у ряда тропических деревьев на стволах и ветвях. Далее они растут до почвы. У ее поверхности они сильно ветвятся и как бы подпирают растение. Пример растения с корнями-подпорками — это баньян.
Воздушные корни
Воздушные корни характерны для орхидей, которые растут на вервях тропических деревьев. Здесь корни орхидей просто свисают вниз.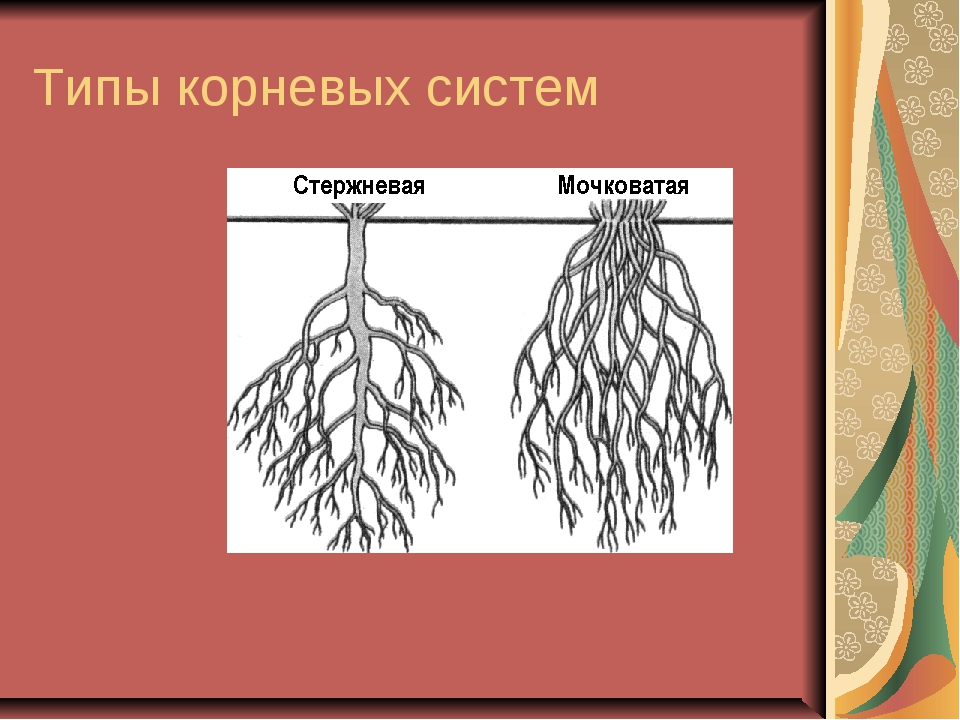 В тропических лесах очень влажно, так что воду можно впитывать прямо из воздуха.
В тропических лесах очень влажно, так что воду можно впитывать прямо из воздуха.
Корни-присоски
Корни-присоски характерны для растений-паразитов. С помощью таких корней растение внедряется в органы других растений и поглощает оттуда воду, минеральные и органические вещества. Такими растениями-паразитами являются повилика и заразиха.
Корни-присоски также можно наблюдать у омелы, погремка, ивана-да-марьи. Они всасывают только воду и минеральные вещества.
Зоны корня
☰
В строении корней большинства растений выделяются несколько зон (перечислены от кончика корня):
корневой чехлик,
зона деления,
зона роста,
зона всасывания,
зона проведения.
Для каждой зоны характерны свои группы тканей и свои функции.
Корень постоянно растет своей верхушкой (кончиком).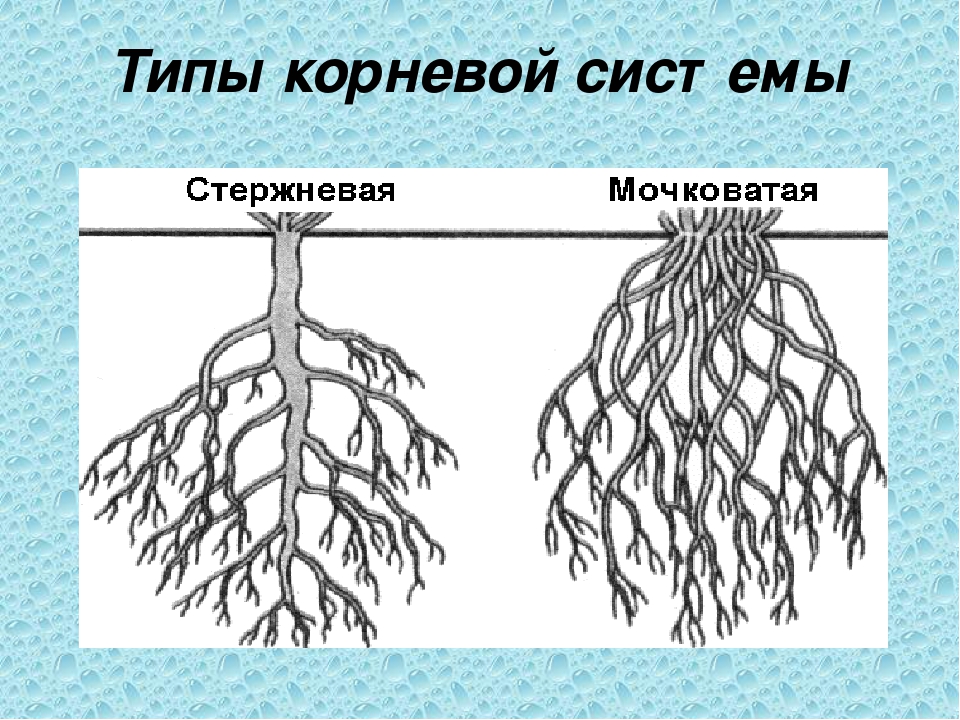 Поэтому клетки одной зоны постепенно превращаются в клетки другой, находящейся дальше от кончика корня (за исключением корневого чехлика). Так, верхние клетки зоны деления становятся клетками зоны роста, а более дальние от кончика клетки зоны роста становится клетками зоны всасывания, клетки зоны всасывания рано или поздно становятся клетками зоны проведения.
Поэтому клетки одной зоны постепенно превращаются в клетки другой, находящейся дальше от кончика корня (за исключением корневого чехлика). Так, верхние клетки зоны деления становятся клетками зоны роста, а более дальние от кончика клетки зоны роста становится клетками зоны всасывания, клетки зоны всасывания рано или поздно становятся клетками зоны проведения.
Корневой чехлик
Корневой чехлик прикрывает кончик корня. У корней многих растений его можно увидеть без увеличительных приборов. Корневой чехлик выглядит как более темное и плотное образование на кончике корня.
Главная функция корневого чехлика — это предохранение верхушки корня, где находится зона деления с клетками образовательной ткани, от повреждений.
Клетки корневого чехлика живые, однако живут мало. Они постепенно слущиваются. От зоны деления образуются новые клетки корневого чехлика.
Те клетки, которые отделяются от чехлика, некоторое время остаются живыми и выделяют слизь, которая облегчает проникновение корня среди частиц почвы, а также растворяет минеральные вещества.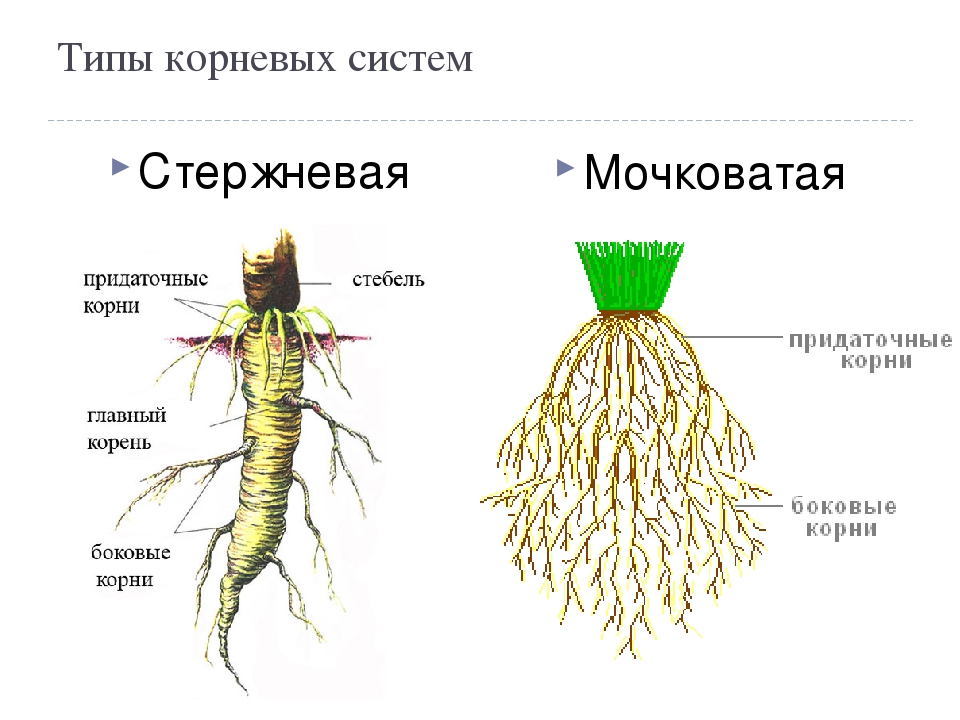
В центре чехлика находятся крахмальные зерна, с их помощью корень определяет, где верх, а где — низ. Корень обладает положительным геотропизмом, т. е. растет вниз.
Зона деления корня
Зона деления находится под корневым чехликом. Ее размер около 1 мм. В этой зоне клетки постоянно делятся.
Клетки зоны деления мелкие, находятся близко друг к другу, их ядра достаточно большие, а цитоплазма густая. Вместе они составляют образовательную ткань.
Зона роста корня
Выше зоны деления находится зона роста корня, составляющая в длину несколько миллиметров. Иногда эту зону называют зоной растяжения. Здесь клетки увеличиваются в размерах, в основном за счет вытягивания в длину. Соответственно, это приводит к росту всего корня в длину. У клеток зоны роста клеточная стенка еще не жесткая, именно это позволяет им растягиваться.
Зона всасывания корня
Зона всасывания находится над зоной роста, обычно ее длина более сантиметра.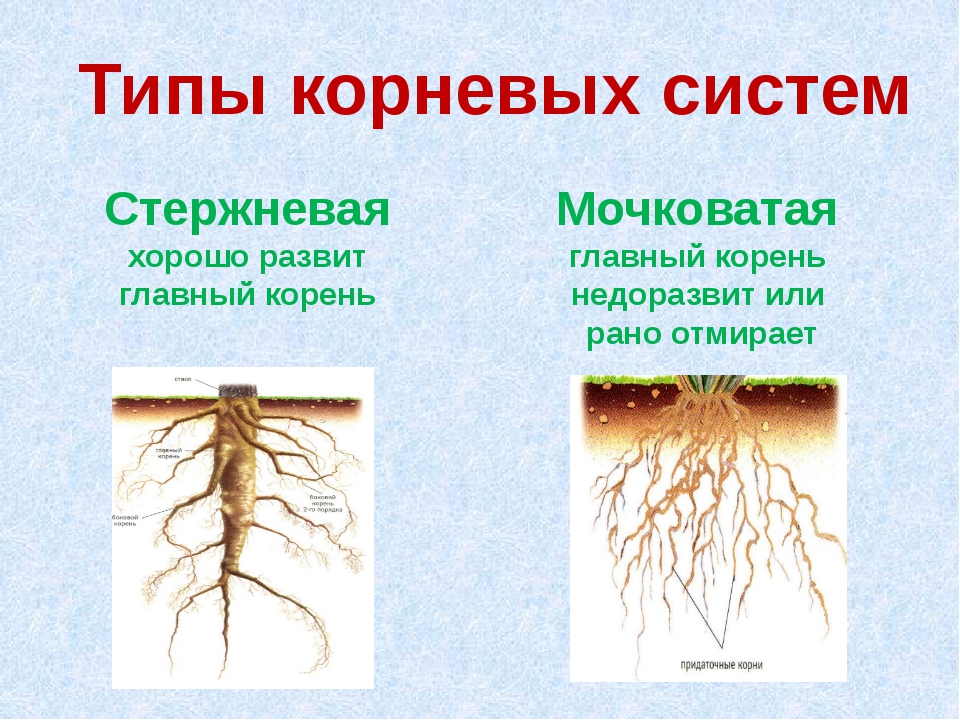 Здесь у каждой поверхностной клетки образуется вырост, который называют корневым волоском. Корневые волоски можно увидеть невооруженным глазом у проростков многих растений. Все вместе они выглядят как пушок, состоящий из беловатых тонких волосков. Каждый волосок обычно в длину не более 1 см.
Здесь у каждой поверхностной клетки образуется вырост, который называют корневым волоском. Корневые волоски можно увидеть невооруженным глазом у проростков многих растений. Все вместе они выглядят как пушок, состоящий из беловатых тонких волосков. Каждый волосок обычно в длину не более 1 см.
Корневой волосок состоит из клеточной оболочки, цитоплазмы, ядра, лейкопластов и вакуоли.
Корневые волоски живут у большинства растений всего несколько дней. Верхние волоски являются более старыми и постепенно отмирают. Зато снизу верхние клетки зоны роста становится клетками зоны проведения. Здесь у поверхностных клеток отрастают волоски.
Главная функция зоны всасывания — это поглощение из почвы воды и растворенных в ней минеральных веществ. Осуществляется эта функция с помощью корневых волосков. Они проникают между частичками почвы, опутывают их и, таким образом, всасывают из почвы водных раствор.
После того как поверхностные клетки всосали водный раствор, он продвигается по внутренним клеткам корня к центральной оси, где находятся клетки зоны проведения.
Зона проведения корня
После зоны всасывания ближе к стеблю находится зона проведения. У этой зоны главная функция — это проведение поглощенного в зоне всасывания водного раствора вверх к стеблю. Водный раствор двигается по сосудам. С другой стороны, от стебля к корню идут органические питательные вещества, корню они нужны для роста, развития и других процессов жизнедеятельности. Органические вещества передвигаются по другим типам клеток.
Волокна проводящей системы есть не только в зоне проведения корня. Ее клетки заходят в другие зоны, расположенные ближе к кончику корня.
Типы корневой системы у растений: примеры с фото
Корневая система
Корневой системой называют все корни растения. Ее формируют главный корень, боковые корни и придаточные корни. Главный корень растения развивается из зародышевого корешка. Придаточные корни обычно отрастают от нижних частей стебля растения. Боковые корни развиваются на главном и придаточном корнях.
Корневая система растений выполняет две основные функции.
Во-первых, она удерживает растение в почве. Во вторых, корни всасывают из почвы необходимые растению воду и растворенные в ней минеральные вещества.
Если у растения развивается мощный главный корень, то формируется стержневая корневая система.
Если же главный корень остается неразвитым или отмирает, а развитие получают придаточные корни, то у растения формируется мочковатая корневая система.
Стержневой тип корневой системы
Для стержневой корневой системы характерен хорошо развитый главный корень.
По внешнему виду он похож на стержень. Главный корень вырастает из зародышевого корешка.
Стержневую корневую систему формирует не только главный корень, но и отходящие от него маленькие боковые корни.
Стержневая корневая система характерна для многих двудольных растений.
Хорошо развитый главный корень есть у фасоли, клевера, подсолнечника, моркови, одуванчика.
Однако у многих многолетних растений с изначальной стержневой корневой системой рано или поздно главный корень отмирает. Вместо него от стебля отрастают многочисленные придаточные корни.
Существует подтип стержневой корневой системы — ветвистая корневая система.
В этом случае сильное развитие получают несколько боковых корней. В то время как главный корень остается укороченным. Тип ветвистой корневой системы характерен для многих деревьев. Такая корневая система позволяет прочно удерживать мощные ствол и крону дерева.
Стержневая корневая система проникает в почву глубже, чем мочковатая.
Мочковатый тип корневой системы
Для мочковатой корневой системы характерно наличие множества приблизительно одинаковых придаточных коней, которые формируют своеобразный пучок.
Придаточные корни отрастают от надземных и подземных частей стебля, реже от листьев.
У растений с мочковатой корневой системой может быть и живой главный корень.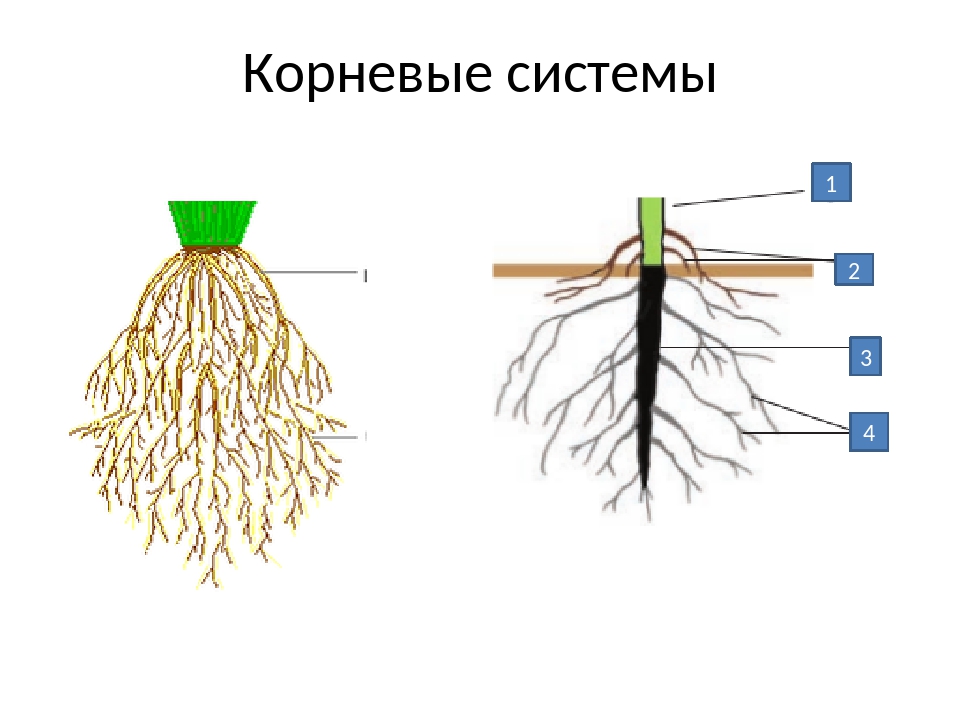 Однако если он сохраняется, то не отличается по размеру от остальных корней.
Однако если он сохраняется, то не отличается по размеру от остальных корней.
Мочковатая корневая система характерна для многих однодольных растений. Среди них пшеница, рож, лук, чеснок, кукуруза, картофель.
Мочковатая корневая система хотя не проникает в почву так глубоко как стержневая, но она занимает большую площадь у поверхности почвы и плотнее оплетает частицы грунта, что улучшает всасывание водного раствора.
Корневые системы и их классификация. Типы корневых систем
Видоизменения корней:
Корнеплод — утолщённый главный корень.
В образовании корнеплода участвуют главный корень и нижняя часть стебля.
Большинство корнеплодных растений двулетние. Корнеплоды состоят в основном из запасающей основной ткани (репа, морковь, петрушка).
Корневые клубни (корневые шишки) образуются в результате утолщения боковых и придаточных корней.
С их помощью растение цветёт быстрее.
Корни-зацепки — своеобразные придаточные корни. При помощи этих корней растение «приклеивается» к любой опоре.
Ходульные корни — выполняют роль опоры.
Досковидные корни представляют собой боковые корни, проходящие у самой поверхности почвы или над ней, образующие треугольные вертикальные выросты, примыкающие к стволу. Характерны для крупных деревьев тропического дождевого леса.
Воздушные корни — боковые корни, растут в надземной части.
Поглощают дождевую воду и кислород из воздуха. Образуются у многих тропических растений в условиях недостатка минеральных солей в почве тропического леса.
Микориза — сожительство корней высших растений с гифами грибов. При таком взаимовыгодном сожительстве, называемом симбиозом, растение получает от гриба воду с растворёнными в ней питательными веществами, а гриб — органические вещества.
Микориза характерна для корней многих высших растений, особенно древесных. Грибные гифы, оплетающие толстые одревесневшие корни деревьев и кустарников, выполняют функции корневых волосков.
Бактериальные клубеньки на корнях высших растений — сожительство высших растений с азотфиксирующими бактериями — представляют собой видоизменённые боковые корни, приспособленные к симбиозу с бактериями.
Бактерии проникают через корневые волоски внутрь молодых корней и вызывают у них образование клубеньков. При таком симбиотическом сожительстве бактерии переводят азот, содержащийся в воздухе, в минеральную форму, доступную для растений.
А растения, в свою очередь, предоставляют бактериям особое местообитание, в котором отсутствует конкуренция с другими видами почвенных бактерий. Бактерии также используют вещества, находящиеся в корнях высшего растения.
Чаще других бактериальные клубеньки образуются на корнях растений семейства Бобовые. В связи с этой особенностью семена бобовых богаты белком, а представителей семейства широко используют в севообороте для обогащения почвы азотом.
Дыхательные корни — у тропических растений — выполняют функцию дополнительного дыхания.
Типы корневых систем
В стержневой корневой системе главный корень сильно развит и хорошо заметен среди других корней (характерно для двудольных).
Разновидность стержневой корневой системы — ветвистая корневая система: состоит из нескольких боковых корней, среди которых не различают главный корень; характерна для деревьев.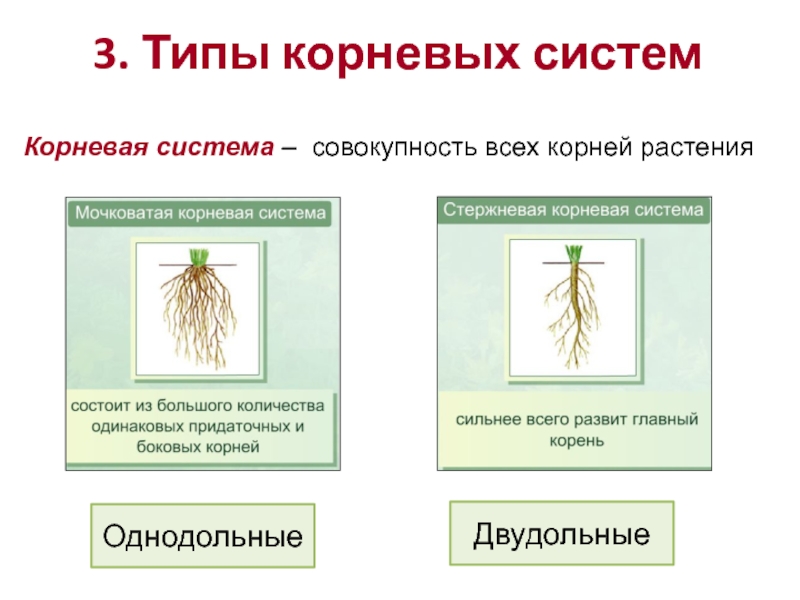
В мочковатой корневой системе на ранних этапах развития главный корень, образованный зародышевым корешком, отмирает, а корневая система составляется придаточными корнями (характерна для однодольных). Стержневая корневая система проникает в почву обычно глубже, чем мочковатая, однако мочковатая корневая система лучше оплетает прилегающие частицы грунта.
Придаточные корни растут непосредственно из стебля.
Они отрастают от луковицы (представляющей собой особый стебель) или от садовых черенков.
Воздушные корни. Корни, которые растут от стебля, но не проникают в землю.
Они используются лазящими растениями для закрепления, как, например, у плюща.
Опорные (ходульные) корни.
Особый тип воздушных корней. Они отрастают от стебля и затем проникают в землю, которая может быть покрыта водой. Они поддерживают тяжёлые растения, например, мангры.
Чем стержневая корневая система отличается от мочковатой?
Корни растения – это его вегетативные органы, находящиеся под землей и проводящие воду и, соответственно, минеральные вещества к остальным, наземным, органам растения – стеблям, листьям, цветкам и плодам.
Но основная функция корня – это все же закрепление растения в грунте.
Об отличительных признаках корневых систем
Общего в разных корневых системах то, что корень всегда делится на главный, боковые и придаточные.
Главный корень, корень первого порядка, всегда вырастает из семени, именно он наиболее мощно развит и растет всегда вертикально вниз.
Боковые же корни отходят от него и называются корнями второго порядка. Они могут ветвиться, и от них отходят придаточные корни, именуемые корнями третьего порядка.
Они (придаточные корешки) никогда не растут на главном, но у некоторых видов растений они могут расти на стеблях и листьях.
Вся эта совокупность корней и называется корневой системой. И существует всего два вида корневых систем – стержневая и мочковатая. И наш главный вопрос касается того, чем же различаются стержневая и мочковатая корневые системы.
Стержневую корневую систему характеризует наличие явно выраженного главного корня, тогда как мочковатая корневая система образована из придаточных и боковых корешков, а ее главный корень не выражен и не выделяется из общей массы.
Чтобы лучше понимать, чем стержневая корневая система отличается от мочковатой, предлагаем рассмотреть наглядную схему строения одной и второй систем.
Стержневую корневую систему имеют такие растения как розы, горох, гречица, валериана, петрушка, морковь, клен, береза, смородина, арбуз.
Мочковая же корневая система есть у пшеницы, овса, ячменя, лука и чеснока, лилии, гладиолуса и других.
Видоизмененные побеги под землей
У многих растения под землей помимо корней имеются так называемые видоизмененные побеги. Это – корневища, столоны, луковицы и клубни.
Корневища растут в основном параллельно к поверхности почвы, они нужны для вегетативного размножения и запасания. Внешне корневище похоже на корень, однако по своему внутреннему строению имеет принципиальные отличия.
Порой такие побеги могут выходить из-под земли и образовывать обычный побег с листьями.
Столонами называются подземные побеги, на конце которых образованы луковицы, клубни и розеточные побеги.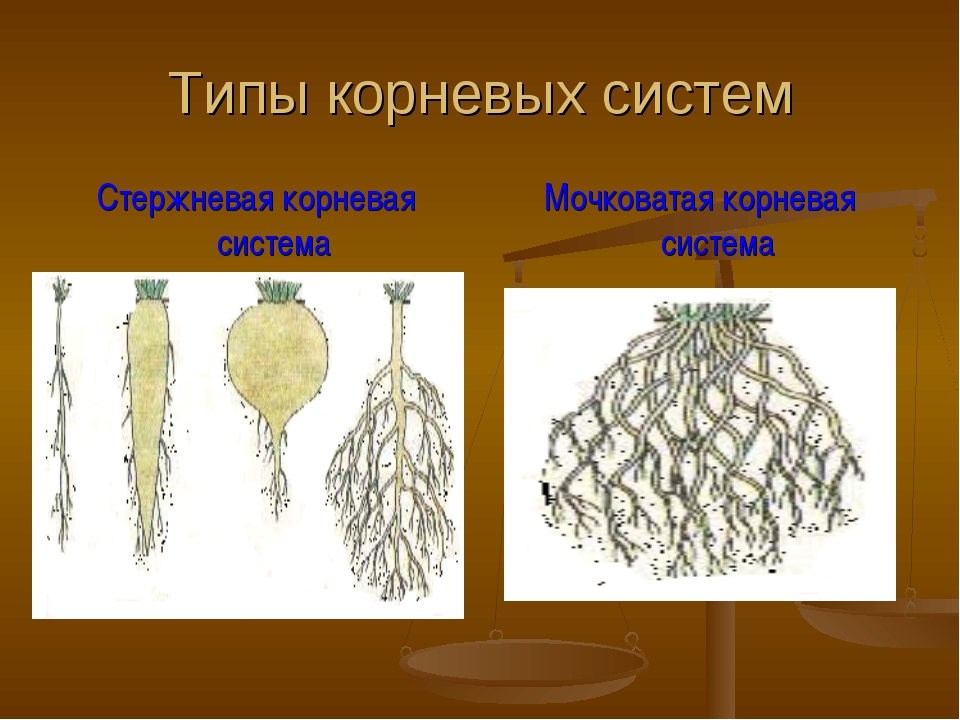
Луковицей называют видоизмененный побег, функцию запасания у которого несут мясистые листья, а от плоского донца внизу отходят придаточные корни.
Клубень – это утолщенный побег с пазушными почками, выполняет функцию запасания и размножения.
Статьи по теме:
| Вейгела — посадка и уход в открытом грунте Декоративные кустарники – давно не редкость в наших садах. И если вы также задумались о приобретении красивоцветущего кустика для своего участка, обратите внимание на вейгелу. О выращивании этого растения в открытом грунте расскажет статья. | Ремонтантная клубника – лучшие сорта Сладкая клубничка хороша и в свежем виде, и для консервирования. Правда, сезон клубники недолог – обычные сорта плодоносят всего пару недель. Чего не скажешь о ремонтантных сортах, радующих урожаем до самых морозов. О лучших сортах ремонтантной клубники – в статье. |
| Безусая ремонтантная земляника — лучшие сорта Грядочки ароматной земляники есть почти на каждом участке. Размножается такая земляника чаще всего усами, но есть и безусые сорта. О них расскажет статья. | Где растут мандарины? Сочные ароматные мандарины – любимый зимний фрукт многих из нас. Даже несмотря на то, что сегодня их можно купить в любое время года, мандарины по-прежнему ассоциируются с новогодними праздниками. Но задумывались ли вы, откуда их привозят? Где растут мандарины – в статье. |
 Особенно популярны ремонтантные ее сорта, дающие урожай по несколько раз за сезон.
Особенно популярны ремонтантные ее сорта, дающие урожай по несколько раз за сезон.Поверхностная корневая система
Поверхностная корневая система образуется у сосны также при неглубоком залегании плотного тяжелого карбонатного суглинка и на таких почвах нередко наблюдается ветровал сосновых семенников, а иногда и семенников лиственницы. Это явление встречается, например, в ряде мест Плесецкого района Архангельской области. На Кольском полуострове ( Мурманская область) ветровал сосновых семенников выражен в местах выхода на дневную поверхлость кристаллических горных пород.
Поверхностная корневая система у сосны, как мы уже говорили, образуется также при неглубоком залегании плотного тяжелого карбонатного суглинка. На таких почвах часто вываливаются ветром сосновые семенники, а иногда и семенники лиственницы, например в некоторых местах Плесецкого района Архангельской области.
На Кольском полуострове ( Мурманская область) и в Северной Карелии ветровал сосновых семенников происходит в местах выхода на дневную поверхность кристаллических горных пород.
Поверхностную корневую систему со слабым развитием вертикально развивающихся корней, всего на 0 5 — 1 м в глубину, образует сосна и на бедных влагой песчаных почвах, где она также может вываливаться сравнительно легко от ветра.
Деревья с поверхностной корневой системой больше подвержены ветровалу, более ослаблены и чаще отмирают на корню.
Диспропорция между усилившейся после рубки транспирацией и ограниченным поступлением влаги из почвы, а также разрывы мелких корней вследствие раскачивания деревьев ветром приводят на неглубоких тяжелых влажных почвах к снижению прироста сразу после рубки. Напротив, деревья на глубоких дренированных почвах, где они образуют глубоко идущие в почву корни и лучше обеспечены влагой, сравнительно хорошо выдерживают изменение обстановки и способны увеличить прирост по диаметру через 2 — 3 года, а иногда и сразу после рубки.
Напротив, деревья на глубоких дренированных почвах, где они образуют глубоко идущие в почву корни и лучше обеспечены влагой, сравнительно хорошо выдерживают изменение обстановки и способны увеличить прирост по диаметру через 2 — 3 года, а иногда и сразу после рубки.
Эти различия отражаются и на анатомическом строении дерева.
Деревья с поверхностной корневой системой больше подвержены ветровалу, более ослаблены и чаще отмирают на корне.
Поврежденная копытами скота поверхностная корневая система ели не в состоянии оказывать сопротивление опенку.
Известны факты воздействия ветра, когда от ветровала страда ПТК с ослабленным дренажом, формирующие поверхностную корневую систему деревьев и расположенные на ветробойных местоположениях.
Ветровал развивается часто в ПТК ельников аккумулятивных склонов с богатыми влажными суглинками, где ель имеет маломощную поверхностную корневую систему. Более ветроустойчивы древостой ПТК денудационных склонов с гльгбово-каменными субстратами, где ель прочно укореняется в расщелинах глыб.
Огонь даже низовой уничтожает тонкокорую, с кроной, низко опускающейся по стволу, с более тонкокорой поверхностной корневой системой ель и пихту и удаляет таким образом сразу два основных препятствия к появлению самосева сосны.
Старые же деревья сосны при любых пожарах имеют шансы уцелеть вследствие более толстой коры, высоко поднятой кроны и весьма глубоко уходящей в почву корневой системы; эти старые деревья остаются в качестве семенников разбросанными в большем или меньшем количестве и после сильных пожаров.
После цветения растения пересаживают в широкие и неглубокие горшки или плошки, так как у азалий поверхностная корневая система, проводят обрезку, удаляя слабые, жирующие побеги и прищипывают верхушки молодых побегов, стимулируя их ветвление. Пинцировку проводят в два-три приема, прищипывая побеги с тремя-четырьмя развитыми листьями. В конце июня пинцировку прекращают, так как в это время на побегах начинается формирование цветочных почек будущего года.
Азалии нуждаются во влажном воздухе.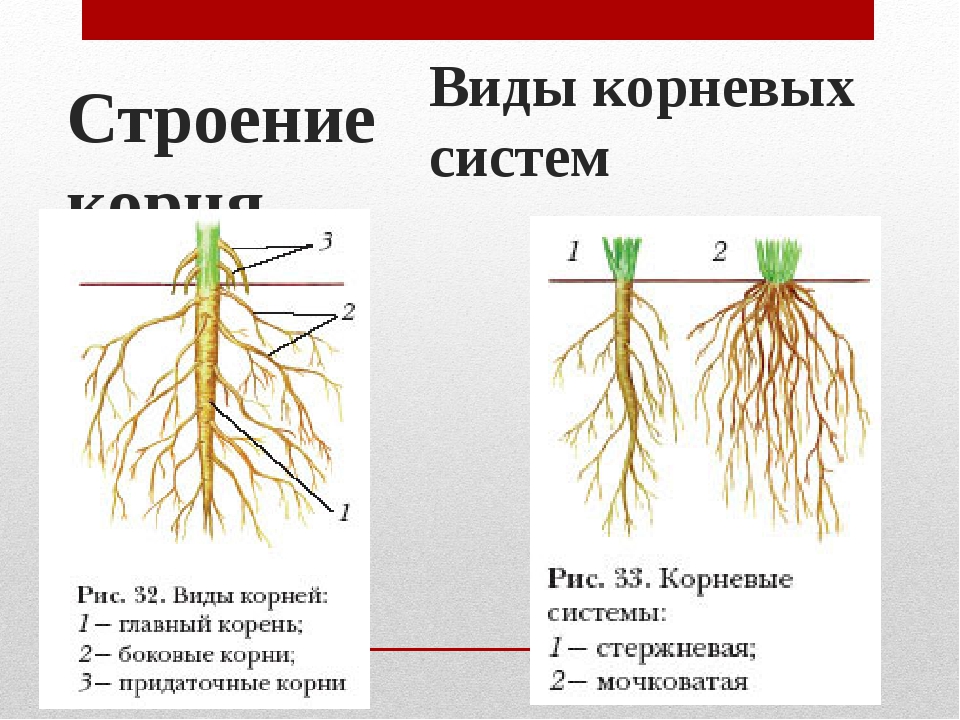 В период активного роста, с марта по сентябрь, их регулярно опрыскивают мягкой водой. Не рекомендуют опрыскивать в период цветения, чтобы избежать появления пятен на цветках. Для нормального цветения нужны высокая интенсивность света и подкормки комплексным удобрением.
В период активного роста, с марта по сентябрь, их регулярно опрыскивают мягкой водой. Не рекомендуют опрыскивать в период цветения, чтобы избежать появления пятен на цветках. Для нормального цветения нужны высокая интенсивность света и подкормки комплексным удобрением.
Веймутова сосна является относительно ветроустойчивой породой, но, как и обыкновенная сосна, может давать и поверхностную корневую систему, например на мелких почвах. К фабричному дыму веймутова сосна не менее чувствительна, чем обыкновенная сосна.
Значительные по площади участки подземных сооружений, обвалованных достаточным слоем земли, озеленяют небольшими группами кустарников, имеющих поверхностную корневую систему, или многолетников.
При необходимости декоративного оформления на них устраивают небольшие рокарии. Во избежание обледенения посадки деревьев и кустарников предусматривают на расстоянии не менее 40 м от открытых брызгальных устройств, а от градирен — на расстоянии не менее 1 5 их высоты.
Корень
Корень выполняет функцию поглощения из почвы воды с минеральными веществами.Он закрепляет и удерживает растение в почве. В корнях могут откладываться запасные питательные вещества.
Строение корня
Корень — осевой орган растения, на котором в отличие от стебля нет листьев. Корень нарастает в длину в течение всей жизни растения, продвигаясь среди твердых частиц почвы. Для защиты нежного кончика корня от механических повреждений и уменьшения трения служит корневой чехлик.
Он образован тонкостенными клетками покровной ткани, которые отслаиваются и образуют слизь, облегчающую продвижение корня в почве. У растущего корня чехлик обновляется каждый день.
Под корневым чехликом располагается зона деления. Она состоит из образовательной ткани.
Клетки этой ткани делятся.
Образовавшиеся клетки растягиваются в продольном направлении и формируют зону растяжения и роста. Это обеспечивает рост корня в длину. Клетки образовательной ткани формируют другие ткани — покровную, проводящую и механическую.
За зоной растяжения следует зона всасывания.
R этой зоне из клеток покровной ткани образуется множество корневых волосков. У пшеницы, например, их до 100 на 1 мм2 поверхности корня. Благодаря корневым волоскам всасывающая поверхность корня увеличивается в десятки и даже сотни раз. Корневые волоски работают как крохотные насосы, которые всасывают из ночвы воду с растворенными в ней минеральными веществами. Зона всасывания подвижна, она меняет свое место в почве в зависимости от роста корня. Корневые волоски живут несколько дней, а затем отмирают, и зона всасывания возникает на вновь вырастающем участке корня.
Поэтому всасывание воды и питательных веществ всегда происходит из нового объема почвы.
На месте прежней зоны всасывания формируется зона проведения. По клеткам этой зоны вверх, в надземные органы, проводятся вода и минеральные вещества, а вниз, от листьев к корням органические.
Клетки покровной ткани зоны проведения у взрослых растений при отмирании могут наслаиваться друг на друга, образуя пробку.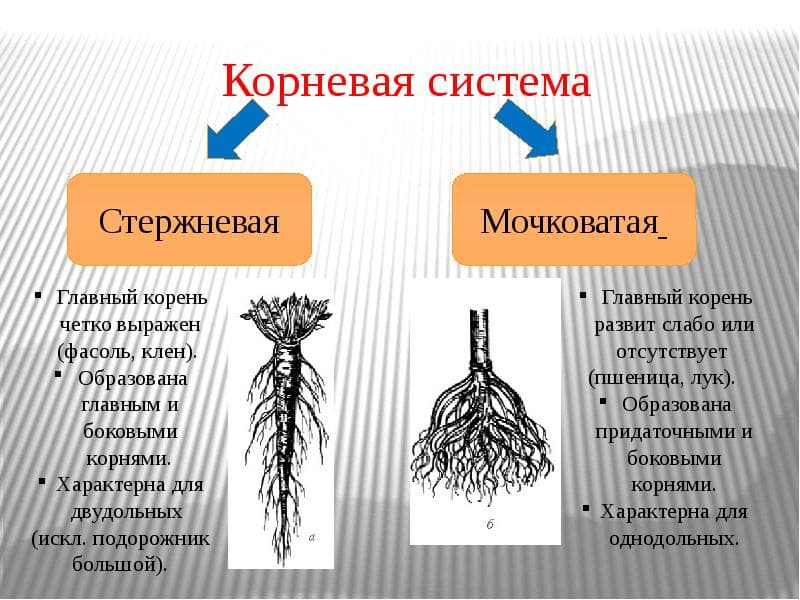 В результате взрослый корень одревесневает.
В результате взрослый корень одревесневает.
На зону проведения приходится большая часть длины долгоживущих корней.
Типы корневых систем
Совокупность всех корней растения называется корневой системой. Различают два вида корневых систем — стержневую и мочковатую.
В стержневой корневой системе выделяют главный корень.
Он растет строго вниз и выделяется среди прочих корней большей длиной и толщиной. От главного корня отходят боковые корни. Стержневая корневая система характерна для гороха, подсолнечника, пастушьей сумки, одуванчика и многих других растений.
Мочковатая корневая система свойственна хлебным злакам, подорожнику и другим растениям, у которых главный корень прекращает рост сразу в начале развития зародыша.
При этом у основания побега образуются многочисленные корни, которые называются придаточными.
Растение развивает пучок, или мочку, более-менее одинаковых по толщине, длине и разветвленности придаточных корней.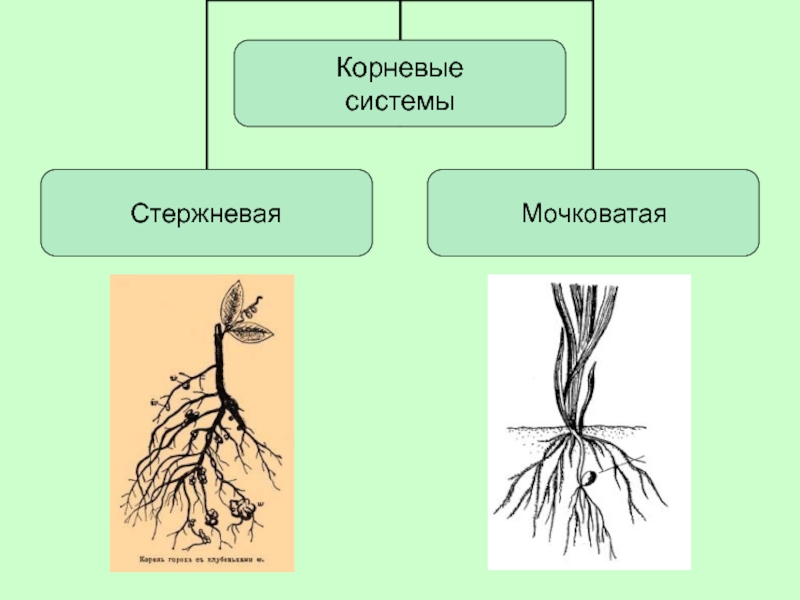
Корень — урок. Биология, Бактерии. Грибы. Растения (5–6 класс).
Функции корня
1. Корни закрепляют растение в почве и прочно удерживают его в течение всей жизни. | 2. Через корни растение получает из почвы воду и растворённые в ней минеральные вещества.
| 3. В корне некоторых растений могут накапливаться запасные вещества. |
Виды корней
Различают три вида корней: главные, придаточные и боковые.
При прорастании семени первым развивается зародышевый корешок. Он превращается в главный корень.
Корни, образующиеся на стеблях, а у некоторых растений и на листьях, называют придаточными.
От главного и придаточных корней отходят боковые корни.
Типы корневых систем
Корни одного растения в почве образуют корневую систему.
Существует \(2\) типа корневых систем:
1. стержневая корневая система состоит из одного главного и множества боковых корней.
стержневая корневая система состоит из одного главного и множества боковых корней.
Пример:
такая корневая система имеется у одуванчика, щавеля, моркови, свёклы и др.
2. Мочковатая корневая система состоит из придаточных и боковых корней приблизительно одного и того же размера. Главный корень у растений с мочковатой системой недостаточно развит или рано отмирает.
Пример:
такая корневая система имеется у пшеницы, ячменя, лука, чеснока и др.
Зоны корня
Кончик корня состоит из мелких живых клеток образовательной ткани. Клетки здесь постоянно делятся, поэтому этот участок корня называется зоной деления.
Кончик корня, как напёрстком, прикрыт корневым чехликом. Корневой чехлик образован клетками покровной ткани, которые защищают нежные клетки зоны деления корня от повреждения твёрдыми частицами почвы. Эти клетки недолговечны, они постепенно отмирают и слущиваются, а взамен отмерших постоянно образуются новые.
Под чехликом расположен конус роста, состоящий из образовательной ткани. Там происходит непрерывное деление клеток. Это зона деления.
За зоной деления находится зона роста (растяжения). Здесь клетки вытягиваются, в результате чего растут в длину.
За зоной роста находится зона всасывания, в которой происходит поглощение воды и минеральных веществ корневыми волосками.
Поглощённая вода и минеральные вещества двигаются вверх по корню по зоне проведения.Корневые волоски
Корневой волосок — это вырост клетки корня.
Большое количество корневых волосков увеличивает поверхность всасывания. Поэтому при пересадке растений корни надо беречь и перемещать с наиболее возможным количеством окружающей их почвы. | Корневые волоски под электронным микроскопом. |
 Корневые волоски непосредственно соприкасаются с почвой и поглощают воду и растворённые в ней минеральные вещества.
Корневые волоски непосредственно соприкасаются с почвой и поглощают воду и растворённые в ней минеральные вещества.
Источники:
Пасечник В. В. Биология. 6 класс // ДРОФА.
http://all-nature.org/korni-rasteniy/
http://fullbiology.ucoz.ru/index/botanika_organy_cv_rastenij/0-293
корневых систем и их типов — StudiousGuy
Все мы знаем, насколько важны корни для выживания растения. Хотя иногда мы не можем их видеть, однако они всегда делают свою работу.
Корни — это не только одна из важных структур, присутствующих в растениях, но они также выполняют различные важные функции. Они играют важную роль в жизненных процессах. Интересно, что не у всех растений есть корни, у некоторых растений, таких как мхи и печеночники, отсутствует корневая система.
Указатель статей (Нажмите, чтобы перейти)
Типы корневой системы
Все корни имеют почти одинаковые функции, но различаются по структуре и месту происхождения.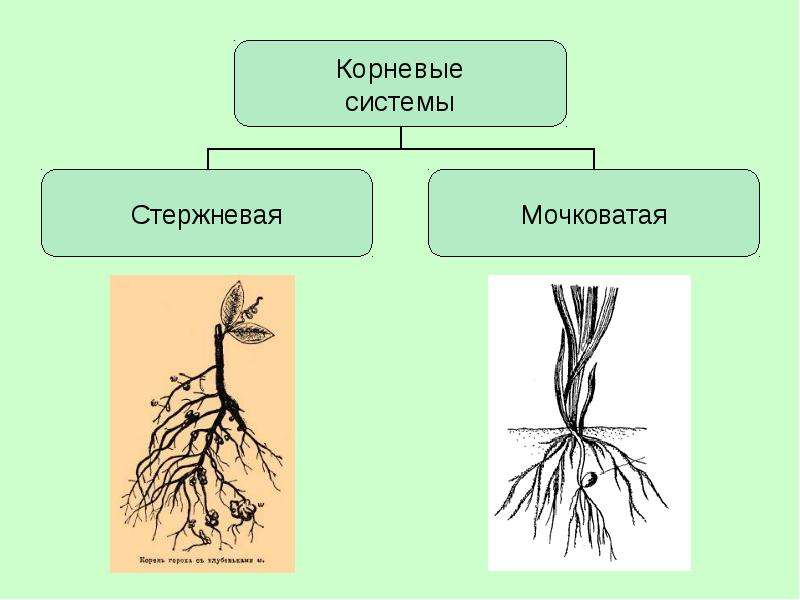 Итак, исходя из этих критериев, корни в основном делятся на три типа, то есть
Итак, исходя из этих критериев, корни в основном делятся на три типа, то есть
Рассмотрим подробнее эти корневые системы.
Система Taproot
Стержневые корни появляются из корешка зародыша прорастающего семени.В системе стержневого корня есть один главный корень, от которого берут начало боковые ветви корня.
При прорастании первый корень, который выходит из корешка, является главным цилиндрическим корнем, также известным как первичный корень, который растет вертикально вниз. Из главного корня возникают корни первого порядка, известные как вторичные корни. А затем из этих вторичных корней возникают корни второго порядка, известные как третичные корни. На кончике третичных корней присутствуют тонкие, похожие на волосы структуры, известные как «корни».«Первичный корень — самый выдающийся, самый толстый и длинный из всех. Стержневой корень постепенно сужается к кончику.
Стержневые корни присутствуют у двудольных растений. Стержневые корни могут сохраняться в течение всей жизни, но на более поздних стадиях развития растения часто заменяются волокнистой корневой системой, потому что корешок большинства видов растений отмирает после прорастания.
Стержневые корни могут сохраняться в течение всей жизни, но на более поздних стадиях развития растения часто заменяются волокнистой корневой системой, потому что корешок большинства видов растений отмирает после прорастания.
Стержневой корень, как правило, глубоко укоренен, и его трудно искоренить. Если в процессе выкорчевывания стержневого растения корень отламывается, оставшаяся в почве часть часто снова дает ростки.
Стержневые корни растут в акропетальной последовательности, то есть самый молодой присутствует на растущем конце, а самый старый — у основания. Корнеплоды — это очень нежные структуры, являющиеся конечными ветвями основной корневой системы. Они отвечают за водопоглощение растений. Первичные корни всегда растут в вертикальной ориентации, а вторичные и третичные корни растут в горизонтальной ориентации.
Примеры системы стержневого корня можно увидеть на множестве растений, даже на тех, которые присутствуют в огородах, например, ;
Дополнительная корневая система
Придаточная корневая система противоположна системе стержневого корня. Придаточные корни растут из других частей растения, кроме корешка или его ветвей. И вместо того, чтобы иметь один первичный корень, из которого возникают ветви, в придаточных корнях существует множество морфологически похожих корней, возникающих из одного и того же узла.
Придаточные корни растут из других частей растения, кроме корешка или его ветвей. И вместо того, чтобы иметь один первичный корень, из которого возникают ветви, в придаточных корнях существует множество морфологически похожих корней, возникающих из одного и того же узла.
Придаточные корни обычно растут над надземными частями растений. Они могут возникать из поврежденных корней, стебля, междоузлий, ветвей или любой другой ткани. Они образуются из предварительно индуцированных первичных клеток корня.
Придаточные корни обычно встречаются у однодольных , но также и у некоторых двудольных, таких как плющ, клубника, клевер и ива. Большинство воздушных корней придаточные. Придаточные воздушные корни многих тропических деревьев вполне обычны. Например,
- Фикус дает свободно свисающие придаточные корни.
- Корни Лианы также происходят от надземных органов.
Придаточные корни могут расти из листовых и стеблевых черенков при помещении в почву. Вы можете просто заново вырастить растения с придаточными корнями вместе с листьями или стеблевыми черенками. Гормоны также вызывают развитие придаточных корней.
Вы можете просто заново вырастить растения с придаточными корнями вместе с листьями или стеблевыми черенками. Гормоны также вызывают развитие придаточных корней.
Примеры придаточных корней:
- Свисающие корни баньяна придают ему механическую прочность.
- Корни мангровых деревьев обеспечивают им устойчивую опору. Они возникают из ветвей и врастают в почву.
- Вьющиеся корни бетеля помогают им крепко держаться за структуру, по которой они поднимаются.
- Опорные корни фиговых деревьев отходят от основания стебля и поддерживают растение.
Волокнистая корневая система
Волокнистые корни возникают из основания стержня или узлов горизонтального стержня; например; травяные растения.
Волокнистые корни не проникают глубоко в почву. Они начинаются как один главный корень, как стержневая корневая система, но позже распространяются, как циновка, по земле, лишь несколько корней уходят глубоко в почву.
В отличие от придаточных воздушных корней, волокнистые корни находятся под землей. Листья с параллельным жилкованием имеют мочковатую корневую систему.
Примеры растений с мочковатыми корнями;
- Просо (Pennisetum glaucum)
- Сладкий картофель (Ipomoea batatas)
- Спаржа (Asparagus officinalis)
Давайте теперь узнаем о некоторых основных функциях корневой системы;
Функции корневой системы
- Корни поглощают воду и минералы из почвы и транспортируют их к верхним частям, таким как стебель и листья растения.Вода и минералы переносятся от корней к верхним частям растений восходящим движением против силы тяжести. Следовательно, требуется некоторая сила. Эта сила обеспечивается транспирационным усилием из-за процесса транспирации, происходящего в листьях.


- Корни также обеспечивают опору для растений и удерживают их прикрепленными к почве, связывая почву вокруг корней. Это причина того, почему сложно выкорчевать растение руками.
- Корни используются для хранения продуктов ; например, мясистые корни, такие как морковь, редис, свекла, репа и мирабилис.
- Корни играют важную роль в связывании почвы. Корни помогают предотвратить эрозию почвы и оползание почвы в холмистой местности.
- Корни таких растений, как клубнелуковицы, стеблевые клубни, корневища и столон, участвуют в вегетативном воспроизводстве.
- Через несколько недель у стебельчатых растений корни помогают цепляться и, следовательно, вьющимся корням; например, Money Plant, Betel, Ivy и др.
- Корни также участвуют в азотфиксации с помощью азотфиксирующих бактерий, присутствующих в почве. Например, узловатые корни гороха, бобов, грамм, пажитника и др.
 Например, узловатые корни гороха, бобов, грамм, пажитника и др.
Например, узловатые корни гороха, бобов, грамм, пажитника и др.- Корни некоторых растений находятся над землей и известны как Воздушные корни . Например, пневматофоры — это тип специализированных воздушных корней, присутствующих у растений мангрового дерева. Они помогают растению дышать.
- Корни мангровых и эпифитных орхидей играют важную роль в фотосинтезе .Под воздействием света они становятся зелеными и начинают вырабатывать энергию. Есть даже орхидея (Dendrophylax lindenii) без листьев, производство энергии которой зависит исключительно от корней.
типов, модификаций и примеров — StudiousGuy
Волокнистые корни — это корни, которые возникают из основания стебля или узлов горизонтального стебля (например, у травы).
Волокнистые корни присутствуют у однодольных растений .
Подобно системе стержневого корня, волокнистые корни также начинаются как один главный корень. Эти первичные корни недолговечны, и поэтому по мере развития растения эти корни заменяются большим количеством волокнистых корней, выходящих из основания стебля. Волокнистые корни распространяются в почве, как циновка; например, корни пальм.
Эти первичные корни недолговечны, и поэтому по мере развития растения эти корни заменяются большим количеством волокнистых корней, выходящих из основания стебля. Волокнистые корни распространяются в почве, как циновка; например, корни пальм.
Корни мочковатой корневой системы морфологически схожи, в отличие от корней в системе стержневых корней, в которых тонкий короткий корень возникает из одного толстого корня.
В этом уроке мы собираемся обсудить различные модификации волокнистой корневой системы и то, как они полезны для растений, а также соответствующие примеры. В конце концов, мы узнаем о некоторых основных функциях этих волокнистых корней.
Указатель статей (Нажмите, чтобы перейти)
Типы фиброзной корневой системы
1. Мясисто-волокнистые корни
Подобно накопительным корням системы стержневого корня, мясистые корни волокнистой корневой системы также накапливают пищу и становятся мясистыми и набухшими.
Клетки волокнистой корневой системы модифицируются таким образом, что они могут накапливать пищу, приготовленную растениями, и хранить ее. Как только пища упакована, эти клетки набухают, и, в конечном итоге, опухает весь корень.
Накопленный корм позже используется растениями в качестве источника питания в неблагоприятных условиях.
Мясистая корневая система делится на три типа, то есть
а. Клубневые или однокорневые клубни
Эти корни встречаются по отдельности, и имеют вид стержневого корня.
Эти опухшие корни не приобретают какой-либо определенной формы. Клубневые корни или запасные корни — это увеличенные боковые корни. Клетки этих корней модифицируются таким образом, что могут хранить в себе пищу.
Пример
- Сладкий картофель ( Ipomoea batatas )
г. Фаскикулированные корни
Это клубневые корни, которые присутствуют в кластерах .
Как и клубневые корни, клетки фасцикулированных корней видоизменены таким образом, что они могут хранить в себе пищу.
Таким образом, единственная разница между клубневыми корнями и пучковидными корнями состоит в том, что клубневые корни появляются поодиночке, а пучковые корни появляются группами.
Примеры
- Спаржа (Asparagus officinalis)
г. Аннулированные корни
Это опухшие корни, имеющие кольцевидную структуру по всей поверхности. Это похоже на диски, поставленные один над другим.Поэтому они называются аннулированными.
Пример
- Ипекак (Цефаэлис ипекакуана)
2. стержневые корни
Эти корни короткие, но толстые, отходят от базального узла стебля. Они растут вниз в косом направлении и проникают в почву. Как только они укореняются в почве, у них развиваются волокнистые корни, которые прочно удерживают почву, поддерживая длинный и узкий неразветвленный стебель, аналогичный веревкам, удерживающим шест или палатку.
Как только они укореняются в почве, у них развиваются волокнистые корни, которые прочно удерживают почву, поддерживая длинный и узкий неразветвленный стебель, аналогичный веревкам, удерживающим шест или палатку.
Эти корни лучше впитывают минеральную соль и воду.
Примеры
- Сахарный тростник (Saccharum officinarum), Кукуруза (Zea mays), Pennisetum и Sorghum (Sorghum bicolor) — их ходульные корни растут в мутовках.
- Винт (Pandanus odoratissimus) — их ходулевые корни отходят от нижней поверхности наклонной ножки и служат опорой.
3. Репродуктивные корни
Эти волокнистые корни мясистые и образуют бутоны. Эти почки при благоприятных условиях превращаются в новые растения. Следовательно, корни, содержащие почки, которые помогают в формировании нового растения, известны как репродуктивные корни.
Примеры
- Сладкий картофель ( Ipomoea batatas )
Функции системы фиброзных корней
- Они поглощают воду и растворенный минерал из почвы.
- В них можно хранить продукты в виде резервного материала.
- Они также могут синтезировать регуляторы роста .
- Они надежно удерживают растение на почве, то есть обеспечивают опору .
типов, модификаций и примеров — StudiousGuy
Придаточные корни — это корни, которые образуются не из корешка, а из других частей растения.Эти корни могут возникать из поврежденного корня, узлов стебля, междоузлий, ветвей или любой другой ткани. Масса придаточных корней вместе со своими ветвями составляет « придаточную корневую систему ».
Придаточная корневая система отличается от системы стержневого корня тем, что вместо одного первичного корня, из которого возникают ветви, в придаточных корнях из одного и того же узла возникают многочисленные морфологически похожие корни.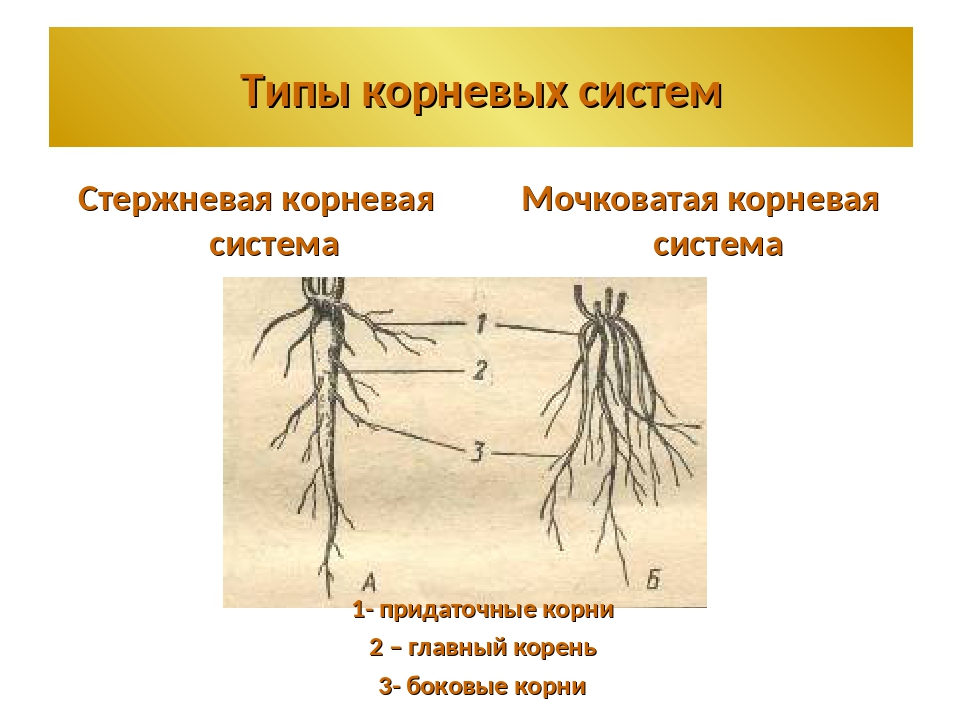
Придаточные корни обычно растут над надземными частями растений.Придаточные корни могут вырасти из листовых и стеблевых черенков при помещении в почву.
Они образуются из первичных клеток корня и обнаруживаются у однодольных растений.
В этом уроке мы узнаем о типах (модификациях) придаточной корневой системы на подходящих примерах.
Указатель статей (Нажмите, чтобы перейти)
Модификации адвентивной корневой системы
Придаточные корни модифицированы для механической поддержки, лазания, , цепляния, и выполнения других жизненно важных функций .Придаточные корни видоизменяются следующим образом:
- Магазин продуктов
- Обеспечить поддержку
- Выполнение дополнительных функций
1. Хранение продуктов питания
A. Клубневые корни
 Клубневые корни
Клубневые корни Клубневые корни являются модификацией придаточных корней. Они мясистые, не имеют особой формы и часто опухают. В случае клубневых корней побеги прорастают с одного конца, а корни — с другого.
Пример — Сладкий картофель (Ipomoea batatas)
B. Фассулированные корни
Фассулированные корни имеют форму гроздей. Эти кластеры возникают у основания стебля.
Пример- Георгин
C. Однородные корни
Еще одна модификация придаточных корней — монилиформные корни. Эти корни тоже опухшие.Однако набухание у однотонных корней происходит через регулярные промежутки времени, что придает корням вид бусинок.
Пример — Розовый мох (Portulaca grandiflora)
D. Аннулированные корни
У этого вида модифицированных придаточных корней на теле присутствует серия выростов, напоминающих форму колец. Последовательность этих наростов выглядит как наложенные друг на друга диски.
Последовательность этих наростов выглядит как наложенные друг на друга диски.
Пример — Ipecac ( Cephaelis ipecacuanha)
E.Нодулозные корни
Нодулозные корни — это модификация придаточных корней, которые вздуты на вершине или на концах. Обладают характерной формой.
Пример — Куркума ( Curcuma longa)
2. Механическая прочность
A. Придаточные корни опоры или столба
- Это тип придаточных корней, которые растут вниз от ветвей деревьев. Эти корни приспособлены для поддержки толстых и тяжелых ветвей.
- Воздушные корни гигроскопичны, то есть они впитывают воду (присутствующую в виде влаги) из воздуха. Из-за поглощения влаги эти корни становятся красными.
- Корни опоры имеют корневых шляпок на концах. Когда корни опоры достигают почвы, они становятся толстыми и похожими на столбы. На этом этапе становится трудно отличить ствол от опорных корней.
 На этом этапе становится трудно отличить ствол от опорных корней.
На этом этапе становится трудно отличить ствол от опорных корней.- Интересный факт заключается в том, что даже если ствол умирает, дерево в целом остается живым, потому что опорные корни дерева поддерживают и питают крону.
Примеры:
- Баньян дерево ( Ficus benghalensis)
Большому баньяну, растущему в индийском ботаническом саду Ачарья Джагадиш Чандра Бос, Ховра, почти 250 лет, и он имеет 1775 опорных корней. Главный ствол этого баньянового дерева сгнил много лет назад. Окружность кроны этого дерева составляет около 404 м.
Самый большой образец дерева, занесенный в Книгу рекордов Гиннеса , найден в деревне Тхиммамма Марриману района Анантапур в Андхра-Прадеш, Индия .Он раскинулся на площади 5,2 соток.
- Человек Роща Растение (Ризофора) )
Б. Стержневые корни
 Стержневые корни
Стержневые корни Эти корни отходят наискось от базального узла стебля и затем проникают в почву. Эти корни короткие, но толстые и приспособлены, чтобы поддерживать растение. Проникая в почву, они превращаются в волокнистые корни и помогают впитывать воду и минералы.
Примеры:
- Сахарный тростник (Saccharum officinarum)
- Fountaingrass (Pennisetum setaceum)
- Сорго (Sorghum bicolor)
- Винтовая шпора ( Pandanus odoratissimus0
C. Вьющиеся корни
- Эти корни встречаются у вьющихся растений (растений, лазающих по различным конструкциям).Они представляют собой неабсорбирующие придаточные корни, которые помогают растению оставаться прикрепленным к структуре.
- Вьющиеся корни проникают в трещины и трещины опоры и помогают растению влезть. Эти корни образуют коготь, набухают или выделяют липкий сок из своего кончика, чтобы прочно удерживать опору.
 Эти корни образуют коготь, набухают или выделяют липкий сок из своего кончика, чтобы прочно удерживать опору.
Эти корни образуют коготь, набухают или выделяют липкий сок из своего кончика, чтобы прочно удерживать опору.- У растения ваниль (Vanilla planifolia) из узелка возникает корень в виде усика. Поэтому они известны как усиковые корни.
- Вьющиеся придаточные корни также могут прорастать из каждого узла и разветвляться.Такие типы вьющихся придаточных корней известны как цепляющиеся корни.
Примеры:
- Tecoma ( Tecoma stans) и Betel (Piper betle) : корни возникают из узлов .
- Вьющаяся фигура (Ficus pumila) : корни отходят от междоузлий .
- Плющ (Hedera): корни возникают из узлов и междоузлий.Кончики корней выделяют липкий сок , чтобы прочно удерживать структуру
- Tecoma ( Tecoma stans) : корни образуют когтей для лазания.
D. Корни контрфорса
- Корни-опоры развиваются у основания стебля и помогают поддерживать структурную целостность растения.
- Базальная часть стебля, удлиненная по вертикали, разносится в почве в разные стороны.
- Этот вид модифицированных придаточных корней дает вид досок.
Примеры:
- Дерево Арджуна (Терминалия арджуна)
- Хлопковое дерево (Bombax ceiba)
- Фиговое дерево Моретон-Бэй в Санта-Барбаре, Калифорния ( Ficus macrophylla )
3. Дополнительные функции
А.Ассимиляционные корни
- Эти корни модифицированы для выполнения жизненно важных функций растений.
- Это зеленые корни, которые способны осуществлять фотосинтез благодаря наличию хлоропластов в их клетках.
- Ассимиляционные корни сильно разветвлены, что увеличивает площадь их поверхности, так что они поглощают максимальное количество солнечного света.
Примеры:
- Водяной каштан (Trapa natans ): их корни погружены в воду, как обычные корни.
- Лунное семя (Tinospora cordifolia ): его ассимиляционные корни, образующиеся из узлов стебля, прорастают в сезон дождей и морщатся во время засухи
- Ленточные корни (Taeniophyllum)
Б. Эпифитные или надземные корни
- Это тип придаточных корней, которые присутствуют в эпифитах, то есть на тех растениях, которые живут на других растениях для укрытия и питания.
- Эпифитные корни неправильной формы свисают с поверхности других растений. Эти корни не имеют корневого чехлика, а вместо этого имеют покрытие из мертвой губчатой ткани, известной как velamen .
- Эпифитные корни гигроскопичны по природе, то есть они поглощают воду, присутствующую в виде влаги, из воздуха с помощью веламена.
Эпифитные растения обладают двумя типами корней; для прилипания (или фиксации) и для поглощения минеральных солей и влаги из пыли, скопившейся на коре.
Примеры:
- Дендробиум (Dendrobium aggregatum)
C. Плавающие корни
- Этот вид придаточной модификации корня встречается у водных растений .
- Эти корни возникают из узла горизонтального плавающего стержня.
- Некоторые из этих придаточных корней накапливают воду и раздуваются. Раздутые корни выходят из поверхности воды и помогают растению плавать.
- Они также помогают в газообмене , поэтому их еще называют дыхательными корнями .
Пример:
D. Микоризные корни
- Микоризы относятся к симбиотической ассоциации гриба с высшим растением.
- Микоризные корни представляют собой классический пример взаимной ассоциации между грибком и корнями.
- Питательные вещества из почвы поглощаются грибком.Растения, в свою очередь, поддерживают грибок органической пищей.
Примеры:
E. Сосущие корни или гаустория
- Чтобы поглощать питательные вещества от хозяина, паразиты развивают микроскопические корни, известные как сосущие корни.
- Поскольку они также встречаются в незеленых паразитических растениях, они также известны как гаустория.
- Сосущие корни прорастают из узлов и проникают глубоко в проводящую ткань хозяина для получения питательных веществ.
Примеры
F. Репродуктивные корни
- Репродуктивные корни — это модифицированные придаточные корни, которые помогают в воспроизводстве.
- Такие придаточные корни дают ростки, которые затем развиваются в побеги.
- Корневой черенок можно посадить в почву, из которой вырастет новое растение.
- Репродуктивные корни являются одним из способов вегетативного размножения .
Примеры:
- Сладкий картофель (Ipomoea batatas)
корневых зон
Корневая крышка
Корневой колпачок представляет собой чашевидную, слабо закрепленную массу клеток паренхимы, покрывающую верхушку корня. По мере того, как клетки теряются среди частиц почвы, из меристемы за крышкой добавляются новые. Шляпка — уникальная особенность корней; кончик стебля такой структуры не имеет. Судя по форме, структуре и расположению, его основная функция кажется очевидной: он защищает находящиеся под ним клетки от истирания и помогает корню проникать в почву.Возникает феноменальное количество крышечных клеток, чтобы заменить те, которые изношены и утрачены, когда кончики корней проталкиваются сквозь почву.
Движению способствует слизистая субстанция mucigel , которая вырабатывается клетками корня и эпидермиса. Муцигель
Смазывает корни.
Содержит вещества, подавляющие корни других видов.
Влияет на поглощение ионов.
Привлекает полезные почвенные микроорганизмы.
Приклеивает частицы почвы к корням, тем самым улучшая контакт почвы с растением и облегчая перемещение воды из почвы в растения.
Защищает клетки корней от высыхания.
Клетки корневой крышечки каким-то пока необъяснимым образом воспринимают свет и направляют рост корней в сторону от света. Корневая шляпка также чувствует силу тяжести, на которую корни реагируют ростом вниз, приводя их в контакт с почвой, резервуаром питательных веществ и воды, используемых растениями.Корневая крышка также реагирует на давление, оказываемое частицами почвы.
Зона деления клеток
Апикальная меристема лежит под и позади корневой крышечки и, как апикальная меристема стебля, производит клетки, дающие начало первичному телу растения. В отличие от меристемы стебля, она находится не на самом конце корня; он лежит за корневой крышкой. Между областью активного деления и крышкой находится область, где клетки делятся медленнее, центр покоя .Большинство делений клеток происходит по краям этого центра и приводит к образованию столбцов клеток, расположенных параллельно оси корня. Клетки паренхимы меристемы маленькие, кубовидные, с плотными протопластами, лишенными вакуолей, и с относительно крупными ядрами.
Апикальная меристема корня организует три первичных меристемы : протодерма , которая дает начало эпидермису; прокамбий , продуцирующий ксилему и флоэму; и земной стержень , который производит кору.Сердцевина, которая присутствует в большинстве стеблей и образуется из наземной меристемы, отсутствует у большинства корней двудольных (эвдикотовых), но обнаруживается во многих корнях однодольных.
30.3A: Типы корневых систем и зоны роста
- Последнее обновление
- Сохранить как PDF
- Ключевые моменты
- Ключевые термины
- Типы корневых систем
- Зоны кончика корня
Кончик корня имеет три основные зоны: зону деления клеток, зону растяжения и зону созревания.
Задачи обучения
- Опишите три зоны верхушки корня и суммируйте роль каждой зоны в росте корня
Ключевые моменты
- Кончики корней в конечном итоге образуют два основных типа корневой системы: стержневые и волокнистые корни.
- Растущий кончик корня защищен корневым покровом.
- Внутри кончика корня клетки дифференцируются, активно делятся и увеличиваются в длине в зависимости от того, в какой зоне расположены клетки.
- Делящиеся клетки составляют зону деления клеток прорастающего растения.
- Новообразованный корень увеличивается в размерах в зоне растяжения.
- Дифференцирующиеся клетки составляют зону созревания клеток.
Ключевые термины
- корешок : рудиментарный побег растения, поддерживающий семядоли в семени и от которого корень развивается вниз; корень эмбриона
- меристема : ткань растения, состоящая из тотипотентных клеток, обеспечивающая рост растений
- прорастание : начало вегетации или роста из семян или спор
Типы корневых систем
Есть два основных типа корневых систем.У двудольных растений стержневая корневая система, а у однодольных — мочковатая корневая система, также известная как придаточная корневая система. Система стержневого корня имеет основной корень, который растет вертикально вниз, из которого возникает множество более мелких боковых корней. Одуванчики — распространенный пример; их стержневые корни обычно отламываются, когда эти сорняки вырываются из земли; они могут вырастить еще один побег из оставшегося корня. Система стержневого корня глубоко проникает в почву. В отличие от этого, волокнистая корневая система расположена ближе к поверхности почвы, где она образует плотную сеть корней, которая также помогает предотвратить эрозию почвы (хорошим примером являются газонные травы, а также пшеница, рис и кукуруза).Некоторые растения имеют сочетание стержневых и волокнистых корней. Растения, произрастающие в засушливых районах, часто имеют глубокую корневую систему, тогда как растения, которые растут в районах с обильным количеством воды, скорее всего, имеют более мелкую корневую систему.
Рисунок \ (\ PageIndex {1} \): Основные типы корневых систем : (a) Системы корневых корней имеют главный корень, который растет вниз, в то время как (b) волокнистые корневые системы состоят из множества маленьких корней.Зоны кончика корня
Рост корней начинается с прорастания семян. Когда зародыш растения выходит из семени, корешок зародыша образует корневую систему.Кончик корня защищен корневой крышкой, структурой, характерной только для корней и не похожей на любую другую структуру растения. Корневой покров постоянно заменяется, потому что он легко повреждается, когда корень проталкивается через почву. Верхушку корня можно разделить на три зоны: зону деления клеток, зону растяжения и зону созревания. Зона деления клеток находится ближе всего к верхушке корня и состоит из активно делящихся клеток корневой меристемы, которая содержит недифференцированные клетки прорастающего растения.Зона удлинения — это место, где новообразованные клетки увеличиваются в длине, тем самым удлиняя корень. Начиная с первых корневых волосков, это зона созревания клеток, где корневые клетки дифференцируются на специализированные типы клеток. Все три зоны находятся примерно в первом сантиметре кончика корня.
Рисунок \ (\ PageIndex {1} \): Зоны кончика корня : Продольный вид корня показывает зоны клеточного деления, удлинения и созревания. Деление клеток происходит в апикальной меристеме.границ | Статистический подход к классификации корневой системы
Введение
Эволюция корневой системы тесно связана с заселением растениями наземных экосистем (Kenrick, 2002; Sperry, 2003), где растениям требуются корни для закрепления, воды и получения питательных веществ. Хотя корни являются ключевым органом адаптации растений к изменчивым условиям окружающей среды и, следовательно, для сохранения биоразнообразия (Cornwell and Grubb, 2003), они долгое время игнорировались в биологии растений и агрономии.Однако в последнее время корневым системам уделяется все больше внимания как ключу к «второй зеленой революции» (Lynch, 2007; Gewin, 2010), ведущей к более ресурсоэффективным растениям.
Надлежащая характеристика разнообразия корневой системы важна для различных целей, таких как улучшение сельскохозяйственных культур, прогнозирование изменений в распределении видов в условиях глобальных изменений или исследование функций корней в углеродном цикле. Чтобы уловить разнообразие, необходима схема классификации корневых систем. Серьезным недостатком является то, что на сегодняшний день у нас нет такой схемы, и лишь несколько недавних публикаций пытаются восполнить этот пробел (например,г., Зобель, Вайзель, 2010). Большинство учебников по ботанике не выходят за рамки очень общего различия в развитии стержневых и волокнистых корневых систем, уделяя больше внимания специализированным морфологическим адаптациям (например, гаусториальным корням, запасным корням), которые встречаются у определенных видов (Bresinsky et al., 2008).
Явное предложение классификации корневой системы было представлено Фиттером (1987) на основе корневой топологии. Фиттер (1987) вывел топологический индекс как меру функционального разнообразия в приобретении ресурсов разными видами.
Другой подход к классификации, основанный на происхождении корней в процессе развития (Cannon, 1949), был недавно повторно предложен Зобелем и Вайзелем (2010). Эта схема основана на различии между первичной, вторичной и третичной корневой системой. Первичная система состоит из первичного (полюсного) корня, происходящего от корешка зародыша, базальных корней, происходящих от гипокотиля / мезокотиля, и боковых корней, отходящих от этих осей. Вторичная система построена из побеговых корней и их боковых сторон, а третичная система относится к тонким корням ниже 0.Диаметр 6 мм. Зобель и Вайзель (2010) и Хоххолдингер и др. (2004) продемонстрировали отчетливый генетический контроль этих типов корней. Тем не менее, при рассмотрении всей корневой системы классификация по развитию приводит только к очень общему различию между корневыми системами с преобладанием стержневого корня и побеговыми корневыми системами. Это позволяет различать гоморизию у однодольных и аллоризию у двудольных, имея ограниченную способность улавливать более подробные различия внутри этих групп.
Классификация корневых систем по их геометрической форме была предложена Kutschera и Lichtenegger (1997). Основываясь на обширных наблюдениях in situ выкопанных корневых систем у более чем 1100 видов, они различали 11 основных типов укоренения от шнуровидного (доминирующий вертикальный рост с небольшим боковым расширением) до дискоидального (доминирующая поверхность вблизи бокового расширения).
Классификация древесных видов, сочетающая морфологические и функциональные признаки, была представлена Вахидом (2000), который определил семь классов различных типов корнеплодов.
Эти схемы классификации корневой системы, представленные до сих пор, различаются по признакам, которые они используют для систематизации, и по степени отражения таксономического разнообразия растений. Традиционная систематика растений использовала филогенетические отношения для определения видов как базовой таксономической единицы с отличительными морфологическими атрибутами (например, Simpson, 2005). Фиттер (1987), однако, ставит под сомнение возможность разработки системы классификации корневой системы на основе видов и выступает за более функциональный подход для выявления различных типов корневой системы.
Функциональные классификации использовались в экологии растений для определения групп видов с общими биологическими характеристиками, которые относятся непосредственно к функции, а не к филогении (Lavorel et al., 1997). Поэтому Вестоби и Лейшман (1997) говорят об экологической, а не таксономической классификации. Хотя функциональные / физиологические признаки могут использоваться непосредственно для классификации (например, Wahid, 2000), основная идея состоит в том, что схема классификации имеет функциональное, а не филогенетическое значение.
Идея функциональных типов, а не видов в качестве единицы классификации кажется особенно полезной для применения к корневым системам. Корни морфологически менее дифференцированы по сравнению с побегами. Следовательно, вероятно, что различия между видами в отношении их корневой системы часто скорее непрерывны, чем дискретны. Четкие, прерывистые группы типов укоренения все еще могут быть обнаружены в результате функциональной адаптации растений к разным условиям окружающей среды (например, Schenk and Jackson, 2002).
Функциональные схемы классификации могут быть получены с использованием субъективных, дедуктивных и определяемых данными методов (Gitay and Noble, 1997). Субъективный метод определяет функциональные группы на основе практического опыта. Считается само собой разумеющимся, что функциональные группы существуют в экосистеме и что они могут быть определены индуктивно с использованием атрибутов растений, рассматриваемых как важные группирующие переменные благодаря знаниям и опыту ботанических экспертов. Дедуктивный подход использует априори установленное представление о важных процессах или свойствах в экосистеме и выводит функциональные категории и связанные с ними наборы характеристик из этих предпосылок.Подход, основанный на данных, использует многомерную статистику для поиска кластеров, возникающих из набора атрибутов. Матрица атрибутов классификации для функциональных групп завода часто зависит от целей и контекста исследования. Поэтому для получения обобщенной классификации некоторые авторы предложили определить базовый набор данных, чтобы расширить сопоставимость между отдельными исследованиями (Weiher et al., 1999). Гитей и Ноубл (1997) рекомендовали тестировать уникальность любого результата классификации на повторяемость, используя одни и те же признаки, измеренные в разное время или в разное время, на соответствие, используя разные атрибуты классификации, и на конвергенцию, используя наборы данных, собранные для разных целей.
Цель данного дискуссионного документа — представить функциональную классификацию корневых систем. Мы показываем недостатки сравнительного исследования корней на основе одного признака и предлагаем новый подход на основе данных с использованием многомерной статистики для получения функциональной классификации корневых систем. Мы предполагаем, что такой подход может предоставить эффективные классификаторы для определения разнообразия корневой системы. Основная цель документа — способствовать обмену мнениями по классификации корневой системы и предложить использование многомерных методов как способ получения дополнительных пояснительных преимуществ от доступных наборов корневых данных в рамках функциональной классификации.
Материалы и методы
Схема классификации, которую мы предлагаем в этой статье, сначала вводится с подробным описанием применяемой статистической процедуры. Затем мы представляем наборы данных, которые используем, чтобы проиллюстрировать подход. Наконец, мы предлагаем базовый набор данных для классификации и объясняем, как наш подход способствует его разработке.
Подход к статистической классификации
Основная цель статьи — предложить метод статистической классификации с определением данных.Мы следовали аналогичным подходам, используемым в экологии для получения функциональных типов растений, в частности, работе Gitay and Noble (1997). Наша процедура основана на анализе главных компонентов (PCA) и двухплоскостной проверке атрибутов корневой классификации с последующим кластерным анализом с использованием корневых типов на основе главных компонентов в качестве составных переменных классификации.
PCA — это процедура для преобразования многомерного набора данных различных переменных признаков в несколько некоррелированных переменных (главных компонентов), которые составляют большую часть дисперсии, существующей в исходном наборе данных.PCA полезен, когда можно предположить некоторую избыточность измеряемых переменных, то есть взаимную корреляцию между признаками, которые измеряют один и тот же конструкт. Извлеченные главные компоненты оптимально описывают общее значение отдельных переменных признаков, которые они содержат. Мы использовали процедуру SAS PROC FACTOR для выполнения PCA. Количество главных компонентов определяется на основе критерия Кайзера (Kaiser, 1960), оставляя любой компонент с собственным значением больше 1,00. Также увеличение объясненной доли дисперсии по каждому последующему главному компоненту является показателем того, сколько компонентов оставить.Чтобы улучшить интерпретируемость основных компонентов, можно применять различные преобразования данных. Распространенным способом является использование вращения, то есть линейного преобразования факторного решения. Мы использовали ортогональное вращение варимакса, которое приводит к некоррелированным главным компонентам. Также могут быть полезны другие преобразования переменных для получения главных компонентов, которые лучше удовлетворяют критерию интерпретируемости. Для этого использовалась процедура SAS PROC PRINQUAL.
Важным результатом PCA является графическое представление решения в виде биплотов.Биплоты содержат отдельные переменные (т. Е. Корневые признаки) как векторы и отдельные объекты (т. Е. Виды, сорта) как точки. Длина вектора равна дисперсии соответствующей переменной. Узкий угол между векторами двух переменных указывает на то, что они сильно связаны друг с другом. Начало диаграммы представляет собой среднее значение для каждой переменной. Чем дальше от начала координат находится проекция объекта на вектор переменной, тем больше этот объект отклоняется от среднего значения переменной.Изучение биплотов позволяет решить, какие переменные важны для представления общей дисперсии исследуемого объекта, то есть какие характеристики необходимы для получения полной картины корневой системы. Это дает возможность гарантировать высокую степень соответствия (т. Е. Одинаковый результат классификации получается с разными наборами атрибутов) при последующей кластеризации. Также повторяемость результата классификации может быть обеспечена при предварительном тестировании отношений между отдельными переменными в разные годы или на разных участках.
Основные компоненты как отличительные составные переменные с разными типами укоренения затем используются в кластерном анализе. Кластерный анализ — это дистанционный метод определения общих групп видов. Это было сделано с помощью процедуры SAS PROC CLUSTER. Здесь следует отметить, что при любом статистическом подходе необходимо учитывать возможное влияние различных процедур математической кластеризации на уникальность решения классификации (например, Milligan and Cooper, 1985; Lo Siou et al., 2011). Ботанические экспертные знания необходимы при сравнении результатов различных процедур кластеризации, которые могут не дать одинаковый результат, а также при принятии решения о количестве значимых кластеров. PROC CLUSTER, однако, обеспечивает некоторую статистическую поддержку принятия решений о количестве кластеров, которые необходимо сохранить (критерий кубической кластеризации, псевдо- F , псевдо- t 2 ).
Моделирование архитектуры корневой системы
Чтобы проиллюстрировать подход к классификации с исчерпывающим набором корневых данных, мы использовали всего 288 смоделированных корневых систем.Они охватывают широкий диапазон разнообразия, начиная от мелких дихотомических типов корней, распространяющихся по бокам, и заканчивая типами «елочки» с преобладанием глубоких первичных корней. Некоторые примеры показаны на рисунке 1.
Рисунок 1. Примеры смоделированных корневых систем, соответствующих шести основным кластерам. (A – C) — это системы стержневых корней (одна ось нулевого порядка), различающиеся по вертикальному распределению корней и интенсивности ветвления, (D – F) — это системы корневых корневищ (четыре оси нулевого порядка) с различным вертикальным распределением и интенсивность ветвления. (A), и (B), — это более «елочные» системы, а (D), и (F), — типичные дихотомические системы.
Для моделирования различных корневых архитектур мы использовали модель Leitner et al. (2010a). Эта основанная на Matlab модель использует L-системы (Прусинкевич и Линденмайер, 1990) для построения геометрии разветвленных корней. Основные правила производства корневых осей включают рост корней, ветвление корней и различные типы тропизма. Удлинение корневой оси следует отрицательной экспоненциальной функции, используемой Pagès et al.(2004). Каждая корневая ось определенного порядка дает боковые ветви. Каждая ось разделена на три зоны: базальная и апикальная зоны около основания и верхушки корня, соответственно, где не образуются ветви, и зона ветвления, где создаются новые корни последовательного порядка. В зоне разветвления появляется заданное количество ветвей. Расстояние между ветками определяется правилом роста сечения. Правило позволяет разветвлениям возникать в любой точке вдоль корневой оси, а не только на краях сегмента с фиксированной длиной сегмента.Модель позволяет моделировать различные тропизмы, вызывающие отклонение кончика корня. На уровне корневой системы могут быть созданы как системы основного корня с преобладанием первичных корней, так и системы мочковатых корней с преобладанием побегов с помощью соответствующей начальной L-системы. В случае системы со стержневым корнем начальная строка состоит из единственного корневого кончика корня нулевого порядка. В случае мочковатых корневых систем определяется количество начальных осей N 0 , а также угол их начального роста в пределах радиуса конуса r 0 .Важно отметить, что модель включает определенное стандартное отклонение для каждого параметра, что дает определенную случайность в каждом прогоне моделирования даже для характеристик, которые были установлены пользователем. Например, влияние частиц почвы на направление роста корней косвенно учитывается некоторым случайным изменением направления роста.
Представление реальных корневых архитектур с помощью имитационной модели было первоначально подтверждено Leitner et al. (2010b) с использованием цифровых изображений выкопанных корневых систем нескольких видов растений из серии корневых атласов Кучера (1960).Их исследование подтвердило надлежащую биологическую основу параметра и правил производства, используемых моделью для моделирования корневой системы растений.
Диапазон параметров модели для моделирования различных корневых архитектур представлен в таблице 1 на основе различных литературных источников.
Таблица 1. Параметры и значения, используемые для моделирования различных корневых архитектур .
В нашем исследовании основной целью применения модели было определение ключевых характеристик, формирующих разнообразие корневой системы, и изучение наиболее эффективных классификационных переменных среди различных характеристик, составляющих сложную общую архитектуру корневых систем.Результаты должны также поддержать разработку списка основных данных и разработку целевых стратегий измерения для классификации корневой системы.
Для этого модельного приложения в код были включены некоторые новые инструменты оценки для получения формы и топологических корневых атрибутов для последующей классификации. Параметры, полученные из смоделированных корневых архитектур, были (i) макроскопической формой латерального и вертикального распределения согласно Vrugt et al. (2001), (ii) топологические параметры ветвления согласно Фиттеру (1987) i.е., величина, высота и внешняя длина пути, и (iii) морфологические характеристики средней оси (общая поверхность корня, средний диаметр, длина на порядок корня). Затем эти корневые атрибуты были переданы в метод статистической классификации, описанный в разделе «Подход статистической классификации».
Измеренная морфология корневой системы
Применение схемы классификации к данным измерений было выполнено с использованием двух выборок морфологических признаков из полевых экспериментов. Характеристики корней анализировали по кернам почвы (диаметром 7 см), где корни были отмыты от почвы, а затем количественно определены с помощью анализа изображений.Анализ изображений с помощью WinRhizo проводился в соответствии с процедурой, описанной в Himmelbauer et al. (2004).
Первый набор данных включал морфологические признаки различных видов, используемых в качестве покровных культур в агроэкологических целях. В этом конкретном случае образцы корней в поверхностном слое были взяты для исследования влияния корней на структурные свойства почвы в поверхностной почве (Bodner et al., 2012). В выборку вошли четыре бобовых ( Vicia sativa L., Lathyrus sativus L., Trifolium alexandrinum L., Melilotus officinalis L.) два brassicaceae ( Sinapis alba L., Raphanus sativus var. oleiformis L.), одна боргинацея ( Phacelia tanacetifolium Benth.), Одна linimaceum usitum at. .), один polygonacea ( Fagopyrum esculentum M OENCH .) и один poacea ( Secale cereale L.). Дополнительно были включены смеси двух видов (Смесь 1; Secale cereale L., Trifolium incarnatum L., Vicia villosa R OTH .; Смесь 2: Phacelia tanacetifolium Benth., Sinapis alba L., Vicia sativa L.).
Образцы были взяты с поверхности почвы (глубина почвы 2–7 см) по две подвыборки на делянку (одна проба в ряду растений и одна проба между рядами) в трех повторностях. Хотя в этом исследовании параметры корней были измерены для конкретной цели (влияние на структуру почвы), критерий сходимости (Gitay and Noble, 1997) утверждает, что такая же классификация должна быть получена, если образцы одного и того же вида были бы отобраны для другой цели. включая другие корневые черты.Помимо основных морфологических признаков, полученных с помощью WinRhizo, мы рассчитали коэффициент диаметра корня как атрибут, описывающий расширение бокового корня. Это было сделано путем сравнения диаметра корней в рядах растений и между ними. Соотношение между двумя положениями отбора проб указывает на изменение диаметра между ветвями около основных осей (ряд) и более удаленных боковых сторон (междоусобицы). Кроме того, однородность между разными порядками корней была аппроксимирована диапазоном диаметров корня, полученным с помощью коэффициента вариации длины корня в разных классах диаметров.Малый диапазон диаметров означает равномерное распределение длины корня по всем классам диаметров.
Второй набор данных был получен из образцов корней 12 генотипов зерновых различных видов и сортов в пределах одного вида. Данные были получены для определения различий в корневых системах, имеющих отношение к засухоустойчивости (Nakhforoosh et al., 2012). Они включали семь T. turgidum subsp. durum; сорта (Floradur и SZD3146 из Австрии, Clovis из Франции, Matt из Аризоны, 7060, 7063 и 7094 из CIMMYT, Мексика), два сорта T.monococcum subsp. monococcum из Турции (PI428154, PI428165), два T. turgidum subsp. turanicum (Камут с Ближнего Востока, TRI5254 из Европы) и один T. timopheevii из Грузии (W9).
Четыре реплики керна почвы были взяты из ряда растений. Образцы из трех глубин почвы (10–20 см, 30–40 см, 50–60 см) были проанализированы WinRhizo. В этом случае распределение по глубине рассчитывалось по наклону линейной регрессии плотности длины корня в зависимости отглубины. Помимо морфологических данных, также была измерена и включена в анализ емкость корня (Chloupek, 1977).
Измеренные данные были сначала проанализированы для каждого признака с помощью одномерного дисперсионного анализа с использованием SAS PROC MIXED. В случае повторных измерений на глубинах использовалась смешанная модель с корреляционной структурой AR (1) для повторяющегося фактора (Piepho et al., 2004). Впоследствии для тех признаков со значительными различиями при p <0,05 среднее сравнение было выполнено с помощью двустороннего теста t .После этого был использован многомерный анализ с PCA и кластеризация для классификации корней на основе морфологических атрибутов, как описано выше.
Базовый набор данных корневых признаков
Предложение о предварительном наборе основных данных по корневым признакам было получено на основе обзора литературы. Цель заключалась в оценке текущей доступности корневых данных, которые можно было бы использовать для классификации. Мы не претендовали на всесторонний обзор литературы, а на приблизительный обзор частоты измерения определенных признаков.Поэтому мы ограничились поиском по ключевым словам в базе данных Scopus за последние 10 лет. Количество совпадений в Scopus использовалось как индикатор потенциальной доступности наборов данных для соответствующих признаков. Предварительная иерархия корневых признаков была установлена на основе шкалы наблюдения. Были перечислены различные корневые атрибуты на каждом иерархическом уровне и указаны обычные методы измерения. В рамках нашего подхода к классификации направление вектора признаков на двух графиках показывает, какие признаки важны для отражения общего разнообразия в выборке и, таким образом, поддерживает разработку основного набора данных.На основе нашей имитационной выборки, охватывающей несколько классов корневых атрибутов, мы показываем, какой класс черт важен для основного набора данных в соответствии с нашими результатами.
Результаты и обсуждение
Настоящий дискуссионный документ мотивирован растущей потребностью в понимании разнообразия и функционирования корневой системы в нескольких фундаментальных и прикладных исследовательских дисциплинах. Для этой цели требуется общая схема классификации для определения отличительных свойств корней у разных видов растений.Здесь мы предлагаем процедуру с определением данных для классификации функциональной корневой системы и иллюстрируем подход с выбранными наборами данных.
Одномерный анализ ограничивает сравнительное исследование корней
Сравнительные исследования корней были проведены в области экологии (например, Schenk and Jackson, 2005) и агрономии (например, Kashiwagi et al., 2005). Когда речь идет о больших выборках, решение об использовании корневого параметра для сравнения сильно зависит от ограничений измерения (например, Manschadi et al., 2007; Chloupek et al., 2010). В большинстве случаев неясно, является ли выбранный признак эффективным для выявления различий в выборке и отражает интересующий функциональный паттерн. Преобладающим методом определения различий в выборке является применение одномерного дисперсионного анализа для каждого отдельного признака.
На Рисунке 2 показан пример этой процедуры.
Рис. 2. Анализ дисперсии выбранных морфологических признаков корней из (A) покровной культуры и (B) образца генотипа зерновых. Изменение соседних полосок порядка вида показывает, что анализ одного признака приводит к различным выводам для каждого признака. (Статистическое сравнение средних значений указано строчными буквами. Виды, имеющие общую букву, существенно не отличаются друг от друга при p <0,05).
Образец покровных культур (рис. 2А) содержал виды из разных семейств растений, описанные по морфологическим признакам корней. Виды бобовых были схожи по корневым атрибутам, связанным с диаметром, но различались по площади поверхности корня.У них был более высокий средний диаметр с меньшей разницей между положением отбора проб в ряду и между рядами (соотношение диаметров), и у них было более равномерное распределение длины корня по разным классам диаметров (диапазон диаметров). Напротив, Brassica вида были похожи по общей площади поверхности корней, но различались по параметрам, связанным с диаметром. Сходство между L. usitatissimum и P. tanacetifolia можно было ожидать по площади поверхности, среднему диаметру и диапазону диаметров, в то время как эти два вида явно различались по соотношению диаметров между рядами.
В образце злаков (рис. 2В) сравниваются более близкие виды одного семейства с использованием соотношения корень: побеги, плотности длины корня, среднего диаметра и распределения глубины корня. Экзотические генотипы ( T. monococcum subsp. monococcum, T. timopheevii ) представляли крайности по большинству признаков, в некоторых случаях (плотность длины, распределение по глубине) с заметными отличиями от других генотипов, которые, однако, не были значимыми в целом. случаи. Европейский T. turgidum subsp. durum Генотипы были очень похожи по длине и диаметру, но имели различное соотношение корень: побеги и распределение по глубине. Опять же, их различия с соседними генотипами (например, мексиканские сорта CIMMYT durum) не были явно значимыми. И снова решение об отличительных группах корневых систем в выборке и любая интерпретация возможных причин вряд ли возможны.
При использовании одномерного дисперсионного анализа для сравнения корневой системы из этих примеров можно выявить следующие проблемы:
и.Ранжирование видов зависит от используемого признака. При использовании разных черт окончательная картина остается неясной.
ii. Одномерный анализ неявно утверждает, что черты независимы друг от друга, хотя они могут быть функционально связаны. Поэтому раздельная оценка каждого признака не может обеспечить всестороннее сравнение на уровне корневой системы.
iii. Маловероятно, что одного корневого признака достаточно для определения структурной адаптации или функционального поведения самостоятельно.В большинстве ситуаций для понимания адаптации и функционирования требуется одновременное рассмотрение более чем одной характеристики (например, реакция корня на уплотнение почвы; Bengough et al., 2006).
iv. Результат сравнения отдельных признаков, вероятно, более специфичен для исследования (разные участки, разные цели) по сравнению с многомерными подходами, охватывающими всю корневую систему. Это легко приведет к противоречивым результатам и затруднению взаимного сравнения исследований.
Сравнительное корневое исследование, таким образом, требует подхода, более адаптированного к системе с множеством признаков, которая вряд ли улавливается, ни структурно, ни функционально, отдельным анализом отдельных атрибутов.
Типы корневого доступа на основе основных компонентов
Чтобы различать различные функциональные типы растений, экологические исследования на уровне всего растения объединили морфологические, физиологические, фенологические и репродуктивные признаки (Duckworth et al., 2000). Фиттер (1987) следовал общей идее функциональной классификации и использовал дедуктивный подход, основанный на предположении, что топологические признаки наиболее адекватны для выявления функциональных различий между корневыми системами. Тем не менее не было продемонстрировано, что топологические классификаторы наиболее эффективно отражают разнообразие корневой системы по сравнению с другими корневыми признаками.Чтобы избежать какого-либо априорного решения по ключевым классификаторам, мы выбрали более открытый подход с определением данных с помощью многомерной статистики (например, Kindscher and Wells, 1995; Naeem and Wright, 2003).
Следуя концепции функционального типа растений из экологии, мы определяем типы укоренения, построенные на отличительной комбинации отдельных признаков, которые будут использоваться в качестве классификаторов. Методологически они получаются с помощью PCA, объединяющего отдельные признаки в независимые составные переменные (главные компоненты).Хэтчер (1994) подчеркивает важность интерпретируемости, т. Е. Новые переменные должны быть биологически значимыми конструкциями. В исследовании засухоустойчивости Araus et al. (2007) продемонстрировали это, используя основные компоненты нескольких надземных морфологических признаков в качестве классификаторов для различного регионального происхождения видов пшеницы.
На рис. 3 показаны биплоиды, полученные из трех наших примеров наборов данных. Каждый биплот содержит векторы корневых признаков и расположение отдельных видов (объектов) в соответствии с оценками их основных компонентов.Для лучшей наглядности векторы признаков и объекты для образца моделирования показаны на отдельных биплотах.
Рис. 3. Биплоты, показывающие векторы признаков и расположение отдельных объектов из (A) образца моделирования, (B) образца видов покровных культур и (C) образца генотипа зерновых. Для лучшей визуализации результатов моделирования векторы признаков и объекты показаны на отдельных биплотах (для объяснения корневых признаков см. Раздел Измеренная морфология корневой системы).
Для смоделированных корневых систем первый главный компонент (PC1) захватывает атрибуты, связанные с плотностью, и топологические параметры Fitter, а второй главный компонент (PC2) в основном связан с диаметром и глубиной укоренения. В соответствии с оценками основных компонентов можно выделить шесть различных типов укоренения. Вдоль ПК1 расположены три группы, различающиеся в основном площадью поверхности, мелкой длиной корня и длиной внешнего пути. Все эти параметры отражают общую плотность укоренения независимо от формы или ветвления.Еще две группы с положительным PC1 различались своим расположением вдоль PC2, одна содержала корни с положительным результатом, а другая — с отрицательными значениями PC2. Таким образом, здесь основным отличием было вертикальное распределение корня (длина стержневого корня, распределение по глубине). Последняя группа была расположена в нижнем левом квадранте с отрицательными баллами PC1 и PC2. Эти системы представляют собой корни типа «елочка» небольшого размера с низким разветвлением и ограниченной глубиной проникновения.
В полевой выборке с видами покровных культур разных семейств растений векторы признаков показывают различие между типами укоренения с преобладанием плотности (высокое соотношение корень: побеги, большая поверхность, длина и мелкая длина) и типами укоренения.Преобладают сорта крупного диаметра (с низкой удельной длиной корня). Корневые системы, относящиеся к типу укоренения с преобладанием плотности, расположены в направлении положительного PC1, тогда как системы корней с преобладанием крупного диаметра — в направлении положительного PC2. В квадрантах с правой стороны двумерного графика (положительный PC1) обнаружены L. usitatissmum , P. tanacetifolia , но также S. alba и смесь 1 с преобладанием горчицы и фацелии. Fabaceae. Все виды встречаются в верхнем левом квадранте, в направлении вектора диаметра и в направлении, противоположном (тонкой) длине корня.Корневые системы большого диаметра / низкой плотности, общие для видов Fabaceae , вероятно, связаны с микробно-корневыми взаимодействиями, характерными для этого семейства растений. Предполагается, что большой диаметр корня бобовых культур был обусловлен развитием их корневых симбионтов (Eissenstat, 1992).
В образце генотипа злаков два основных типа укоренения также связаны с высоким (положительный PC1) и большим диаметром / массой (положительный PC2). Здесь также распределение по глубине содержится в PC1, с поверхностью около корневой системы на правой стороне двукратного участка (положительный PC1) и глубоко укоренившимися видами на левой стороне (отрицательный PC1).Это указывает на компромисс между высоким потенциалом эксплуатации ресурсов за счет плотной корневой системы и высокой исследовательской способностью глубоко укоренившихся типов (Fitter et al., 1991; Fitter, 2002). Дикие генотипы ( T. monococcum, subsp. monococcum, T. timopheevii ) расположены в правой части двумерного графика (положительный PC1), особенно в нижнем квадранте. Таким образом, для них характерны корневые системы высокой плотности с поверхностью, близкой к концентрации корневых осей. T. turgidum subsp. durum сорт Matt и T. turgidum subsp. turanicum разновидность Kamut оба расположены в нижнем левом квадранте, разделяя высокое отрицательное значение PC2. Они демонстрируют тип укоренения с плотностью ниже средней и большим количеством корней в глубоких слоях почвы. Все центральноевропейские генотипы расположены в верхнем левом квадранте с высоким PC2 и отрицательным PC1, то есть демонстрируют глубоко укоренившийся тип укоренения с преобладанием диаметра. Генотипы CIMMYT обнаруживаются вокруг происхождения двух участков, что указывает на промежуточный тип укоренения.
Несмотря на различное количество и тип корневых признаков в каждом наборе данных, во всех случаях было сильное влияние признаков плотности укоренения (например, длины, мелкой длины и площади поверхности) на первый главный компонент, в то время как второй Главный компонент всегда был положительно связан с высоким средним диаметром и преобладанием осей грубого корня. Общее значение, лежащее в основе основных компонентов, демонстрируется количественно значительным соотношением оценок основных компонентов общих корневых признаков между различными выборками (моделирование vs.Образец покровной культуры: r 2 = 0,65, p = 0,02 *; моделирование против образца зерновых: r 2 = 0,82, p = 0,002 **; Покровная культура против образца зерновых: r 2 = 0,65, p 0,05 *). Это доказывает, что основные компоненты действительно предоставили значимые конструкции, выражающие различные типы укоренения.
В наших примерах типы укоренения на основе главных компонентов охватывают от 74% (образец покровных культур) до 89% (образец зерновых) от общей изменчивости.Третий главный компонент увеличил бы объясненную дисперсию от 6,6% (смоделированная выборка) до 14,5% (выборка покровных культур) и мог бы быть рассмотрен в некоторых случаях.
Функциональная классификация на основе типов укоренения
Наш подход, основанный на данных, направлен на построение функциональной классификации на основе основных компонентов корневых типов, объединяющих всю доступную информацию о корневых признаках. Каждый вид количественно характеризуется оценками основных компонентов, которые определяют его связь с различными типами укоренения.
Затем выполняется этап классификации с использованием кластерного анализа. Этот статистический метод определяет расстояние между видами, содержащимися в выборке. Полученные дендрограммы иллюстрируют количество функциональных групп, которые появляются из данных, содержащих все виды со схожими типами укоренения. Возникающие группы представляют собой адекватную функциональную классификационную единицу, которую можно использовать для дальнейшей интерпретации основных причин (например, генетическое родство, адаптация к окружающей среде). На рисунке 4 показаны дендрограммы, рассчитанные на основе трех наших примеров наборов данных.
Рис. 4. Дендрограммы, показывающие результат классификации на основе типов корневого доступа на основе главных компонентов, используемых в качестве классификаторов. Результаты взяты из (A), — имитационная выборка, (B) — выборка видов покровных культур и (C) — выборка генотипа зерновых.
Для смоделированной корневой выборки статистические критерии (критерий кубической кластеризации, псевдо- F и псевдо- t 2 ) предлагают шесть групп, которые следует сохранить в дендрограмме.Есть два четко различимых кластера с наибольшим средним расстоянием. Они связаны с количеством осей нулевого порядка (один против четырех корней нулевого порядка). Однако общее количество кластеров с отличительными группами, которые следует учитывать в наборе данных, равно шести. Эти группы различаются (i) количеством осей нулевого порядка, (ii) количеством боковых ветвей и (iii) расстоянием между ветвями между боковыми линиями вдоль оси нулевого порядка. Все три первых кластера сверху вниз вдоль пунктирной линии дендрограммы представляют системы стержневых корней с увеличивающимся числом боковых ветвей.Расстояние между ветвями на стержневом корне одинаково в кластере 1 и 3, тогда как в кластере 2 длина между ветвями примерно в два раза больше. Кластеры с четвертого по шестой содержат системы с четырьмя корнями нулевого порядка (приближение побеговых корневых систем), снова с увеличением числа боковых ветвей с четырех до шести. Расстояние между ветвями корней нулевого порядка является самым высоким в пятом кластере и самым низким в шестом кластере.
Принимая во внимание корневые признаки, содержащиеся в типах укоренения на основе главных компонентов, первый, второй и шестой кластеры в основном различаются плотностью укоренения (оценка PC1), а для кластеров с третьего по пятый основным отличительным критерием является их распределение по глубине ( Оценка PC2).Если рассматривать корневую топологию как классификатор, первый и второй кластеры относятся к категории слаборазветвленных систем типа «елочка», а шестой кластер представляет собой сильно разветвленную дихотомическую систему.
Используя только форму и морфологические индикаторы, можно было бы сохранить четыре отчетливых кластера. Такое же количество кластеров можно было бы получить только с учетом топологических индикаторов. При сведении классификационных переменных только к морфологическим атрибутам с одной осью (диаметр, площадь поверхности, мелкая длина) были бы идентифицированы три отличительные группы (дендрограммы не показаны).
Результаты классификации на основе данных моделирования корня показывают, что тип развития (ветвление от одной оси нулевого порядка, исходящей от зародышевого корешка, по сравнению с ветвлением от нескольких осей нулевого порядка, исходящих от узлов побега) действительно является фундаментальным отличительным критерием при высокой иерархический уровень. Это согласуется с недавней классификацией развития Zobel and Waisel (2010), показывающей решающую роль происхождения корня для понимания различных структур корневой системы.Некоторые морфологические свойства, связанные с плотностью укоренения, сильно зависят от типа развития. Тем не менее, результаты также подчеркивают нашу критику классификаций развития. Они ограничены очень грубым различением (система отстукивания от побегов) и недостаточны для выявления всех существующих групп с отличительными типами укоренения.
Для выделения всех шести отличительных групп, идентифицированных статистическими показателями, необходимы классификаторы, связанные с плотностью, формой (распределение) и топологическими классификаторами.Параметры топологической классификации из Fitter (1987) и морфологические атрибуты с параметрами макроскопической формы являются одинаково эффективными классификаторами при отдельном использовании (четыре кластера). По-прежнему недостатком топологического подхода является сложность измерения топологической ветвящейся структуры. Об этом свидетельствует небольшое количество исследований, в которых представлены данные такого типа (см. Таблицу 2). Однако с быстрым увеличением in situ возможностей построения изображений мы можем ожидать лучших архитектурных данных для топологической классификации в будущем (Zhu et al., 2011; Ingram et al., 2012).
Таблица 2. Признаки в различных масштабах наблюдения для исчерпывающего, иерархически упорядоченного набора основных данных для классификации корневых систем, общих методов измерения и совпадений в базе данных Scopus из поиска по ключевым словам .
Напротив, адекватные схемы пространственной выборки для морфологических признаков (Bengough et al., 2000) для получения параметров формы, которые используются здесь (Vrugt et al., 2001), предоставляют более легкодоступные классификаторы, которые также охватывают большую часть разнообразия, что приводит к значимому классификация.Морфология осей и макроскопическое распределение кажутся разумным компромиссом между грубым различием, обусловленным типом развития, и тонкой дифференциацией, которая требует информации о топологическом ветвлении.
Для набора данных по видам покровных культур четыре группы предложены критерием кубической кластеризации. Дендрограмма четко разделяет бобовые на одном конце, а L. usitatissmum образует отдельную группу на противоположной стороне. По плотности преобладали укореняющиеся типы ( Brassicaceae, , P.tanacetifolia , смесь 1) образуют общую группу, тогда как S. cereale , F. esculentum и смесь бобово-рожь 2 представляют собой промежуточную группу между типами укоренения с преобладанием диаметра и плотности. Различие между системами бобовых культур с преобладанием большого диаметра и системами Brassica с преобладанием плотности можно интерпретировать функционально как связанное либо с сильным эксплуатационным потенциалом (тонкие корни, высокая поверхность контакта корня с почвой), либо с высокой исследовательской способностью (Fitter, 2002), в случае бобовых культур, также подверженных колонизации ризобиями.
По таксономическому порядку семейств растений данные по видам покровных культур показали, что только для определенных семейств можно определить характерный образец укоренения. Fabaceae четко сгруппированы вместе, имея тип укоренения с большим диаметром / низкой плотностью (высокий PC2). Также вида Brassica и вида были в объединенном кластере (тип плотности с положительным PC1). Однако они делили свой кластер с видами из других семейств. Это говорит о том, что либо морфологических дескрипторов недостаточно для выявления различий в корневых системах этих семейств растений, либо действительно эти виды, используемые в качестве покровных культур, выращиваемых осенью, имеют схожие типы укоренения из-за сходной адаптации к окружающей среде, несмотря на принадлежность к разным семействам.
Генотипы злаков делятся на четыре отдельные функциональные группы. Мексиканские генотипы CIMMYT durum сгруппированы на одном конце дерева, а Мэтт и Камут — на другом. Последние образуют низкоплотный тип с глубокими корнями (низкий PC1 и низкий PC2), тогда как генотипы CIMMYT являются промежуточным типом (PC1 и PC2 вблизи происхождения). Отдельную группу составляют T. monococcum subsp. monococcum и T. timopheevii , характеризующиеся типом неглубокого укоренения с преобладанием плотности (высокий ПК1).Четвертый кластер включает среднеевропейские сорта твердых сортов, а также европейский T. turgidum subsp. turanicum Генотип с укоренением большого диаметра, типы средней плотности и глубины (высокий PC2 и PC1 вблизи происхождения).
Набор данных по зерновым показывает связь между региональным происхождением и типом укоренения. Генотипы из летнего засушливого климата (Arizona T. turgidum subsp. durum var. Matt and Middle Eastern T. turgidum subsp. turanicum var. Камут) имеют общий кластер, отличный от всех остальных, несмотря на принадлежность к разным подвидам. Также T. turgidum subsp. turanicum var. TRI5254 с неустановленным европейским происхождением попадает в общий кластер с центральноевропейскими твердыми видами, что, по-видимому, связано с региональным происхождением. В случае экзотических генотипов ( T. timopheevii и T. monococcum subsp. monococcum ), имеющих общий кластер из разных подвидов, вероятно, низкая интенсивность размножения является основной причиной их сходных корневых систем ( Рейнольдс и др., 2007). Их общий рост больше напоминает естественные виды трав, чем современные зерновые культуры. Кластер, образованный сортами CIMMYT durum, можно было ожидать, поскольку они имеют общий генетический фон. Хотя региональное происхождение важно во многих случаях, все же сходство в геноме, по-видимому, является основным фактором, лежащим в основе совместных кластеров: T. turgidum subsp. turanicum и subsp. durum оба имеют геном BA u , а T. monococcum (A m ) и T.timopheevii (GA m ) оба содержат геном A m (происходящий от T. uratu ).
Результаты классификации из двух выборок полей подтверждают, что значимые группы могут быть извлечены на основе типов укоренения на основе главных компонентов. В большинстве случаев можно было выявить возможные причины появления общих групп видов. Отличительные группы показали связь с генетическим фоном, предполагая, что также на уровне корневых систем можно ожидать сходства из-за филогенетической связи.Тем не менее, общая адаптация к окружающей среде и селекционный фон кажутся важными причинами формирования корневой системы. В некоторых случаях более точное различие может быть получено при включении топологических корневых атрибутов. Однако различные потенциальные причины общих кластеров также подчеркивают идею функциональной классификации, в которой движущие силы окружающей среды могут иметь доминирующее влияние на тип укоренения за пределами филогенетических отношений.
Основной набор данных для классификации
Мы показали, что PCA предоставляет значимые и эффективные классификаторы, которые объединяют всю доступную корневую информацию без предварительного определения их важности для сравнительной классификации.Некоторые авторы, работающие над функциональными классификациями, предложили базовые наборы данных для улучшения сопоставимости результатов классификации (Weiher et al., 1999). В оптимальном случае признаки в наборе основных данных о корнях должны соответствовать трем основным критериям: (i) они должны обеспечивать биологически значимое и полное описание корневой системы; (ii) они должны содержать признаки, позволяющие эффективно различать корневые системы и отражать как можно больше деталей видовых различий; (iii) он должен содержать признаки, которые можно легко измерить с помощью согласованных протоколов.
Таблица 2 предлагает список основных черт корня, охватывающий различные структурные и функциональные масштабы корневой системы. Иерархический порядок признаков предполагает, что дискретные классы корневых систем можно найти на более высоком уровне (общий размер и форма корневой системы, тип развития), тогда как на более низком уровне (морфология отдельных осей, физиологическое функционирование) различия более продолжительны. Однако из этого правила могут существовать важные исключения (например, сильно отличительные анатомические признаки).Мы также предоставляем соответствующие методы измерения и наличие признаков через количество совпадений при поиске в базе данных Scopus. Обзор базы данных ясно показывает, что информация о корнях в основном доступна на уровне общей биомассы корней (27% исследований). Также чаще сообщалось о морфологии одиночных осей (8%) и некоторых физиологических признаках корня (например, дыхание, аквапорины; 6%). Напротив, архитектурные черты редко измеряются, особенно в зрелых корневых системах, и некоторые соответствующие физиологические параметры (например,g., корневые сопротивления) вообще отсутствовали в нашем скрининге базы данных.
Райх (1993) предполагает, что количество признаков, необходимых для минимального набора данных, можно уменьшить с помощью статистических методов, исследующих ковариацию между признаками. В рамках нашего подхода к классификации, основанной на данных, это можно сделать до кластеризации во время проверки двух участков. Когда векторы признаков имеют одинаковое направление, они демонстрируют совместную изменчивость. В этом случае одни дескрипторы могут быть заменены другими (например, более легко измеряемыми признаками) без потери информации обо всем разнообразии системы и обеспечения согласованности последующего результата классификации (см.Приложение). Кроме того, стабильность признака и, следовательно, ожидаемая повторяемость результата классификации могут быть оценены с помощью двухплоскостной проверки в случае множественных местоположений или многолетних наборов данных. В то время как, например, диаметр корня является довольно стабильным признаком, распределение корней и соотношение корней и побегов сильно зависят от различных условий окружающей среды (см. Приложение). Для признаков, сильно реагирующих на различные условия окружающей среды, можно включить индекс пластичности (например, Bell and Sultan, 1999; Valladares et al., 2006), чтобы зафиксировать влияние взаимодействия корня с почвой на структуру корневой системы.
Также модели корневой архитектуры могут быть важным инструментом для поддержки разработки списка основных характеристик для классификации. Насколько нам известно, после публикации Fitter (1987) и его последовательных исследований, мы первыми применили модель корневой архитектуры в контексте классификации. Основное намерение состояло в том, чтобы определить сильные классификаторы, чтобы различать различные корневые системы в пределах архитектурного разнообразия, ожидаемого в природе. Наши результаты показывают, что информация о морфологии осей, форме корневой системы и ветвлении (топологии) необходима для захвата всех отличительных групп в большой выборке.Отсутствие данных архитектурных измерений может лишь частично компенсироваться другими корневыми атрибутами, такими как дескрипторы макроскопической формы.
Результаты классификации, представленные в этом исследовании, были ограничены различиями в структурных характеристиках корня, а не в функциональности (например, поглощение воды или питательных веществ). При включении функциональности другие признаки (например, углы ветвления, тропизмы) могут стать более важными для фиксации отличительных типов укоренения. Соотношение между классификациями на основе структурных истрого функциональные / физиологические атрибуты могут быть изучены теоретически при расчете функциональных различий, возникающих в результате моделирования различных корневых архитектур.
Хотя до сих пор отсутствует определенный тип корневых данных (например, зрелая архитектура корневой системы), экспоненциальный рост исследований корневых систем с 1960-х годов убедительно свидетельствует о том, что общая классификация корневых систем не является в основном проблемой отсутствия данных, а скорее согласованного усилия и метода объединения существующих знаний.Представленная здесь схема классификации с определением открытых данных могла бы внести вклад в устранение этого пробела в корневых исследованиях.
Выводы
В данной статье представлен подход с определением данных для классификации корневых систем. Цель состоит в том, чтобы предоставить основу для определения основных отличительных свойств корневой системы разных видов, проанализировать потенциальные движущие силы в эволюции разнообразия корневой системы и изучить функциональное значение различных стратегий укоренения. Мы следуем концепции «функциональных типов растений», используемой в экологии и экофизиологии, для поиска общих групп на функциональной, а не филогенетической основе.Мы предлагаем подход, основанный на данных, в котором отдельные группы получаются с помощью методов статистического исследования данных без предварительного решения по выбранному классификатору.
Наши основные выводы заключаются в том, что морфологическое описание корня с соответствующими пространственными данными обеспечивает надежные атрибуты для классификации различных типов корневых систем. Это было продемонстрировано как смоделированными, так и полевыми измерениями корневых данных, которые позволили идентифицировать основные распространенные типы укоренения на основе средних морфологических признаков осей.Тем не менее, при интеграции топологической информации можно ожидать более точной классификации. Типы корневой системы являются совместным результатом филогенетических связей и давления окружающей среды, а также человеческого отбора. Таким образом, функциональная классификация является наиболее подходящей для выявления разнообразия корневых систем. Подход, основанный на данных, может объединить все большее количество знаний о корневой структуре и функционировании. Проверка данных на основе PCA и двух графиков предоставляет методы для определения ключевых характеристик для основного набора данных и обеспечивает высокую степень стабильности при группировании на основе кластеров.
Метод классификации с определением данных поощряет объединение результатов из различных шкал измерения и направлений исследований, чтобы охватить общее разнообразие корневой системы для широкой схемы классификации. В настоящее время обширная информация о морфологии и пространственном распределении средних осей доступна в результате эффективных инструментов анализа изображений. Наше исследование предполагает, что такие наборы морфологических данных (длина, диаметр, распределение по глубине) будут представлять собой надежный начальный шаг для классификации за пределами установленного грубого различия, основанного на типе развития.Однако для подробной классификации корневых функциональных типов базовый набор данных требует количественных знаний о корневом ветвлении. Будущая интеграция корневой функциональности еще больше расширит основу нашего подхода и подчеркнет роль функциональных и структурных атрибутов для классификации корневых функциональных типов.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Исследование выполнено в рамках проекта «Корни засухоустойчивости. Понимание экогидрологии типов корневой системы староместных сортов твердой пшеницы для повышения устойчивости к засухе ». Мы признательны за финансовую поддержку Австрийскому научному фонду (FWF): номер проекта P 25190-B16. Даниэль Лейтнер является стипендиатом APART Австрийской академии наук в Центре вычислительных наук Венского университета.
Сноски
Список литературы
Араус, Дж.Л., Феррио, Дж. П., Буксо, Р., и Вольтас, Дж. (2007). Историческая перспектива земледелия засушливых земель: уроки, извлеченные из 10000 лет выращивания пшеницы. J. Exp. Бот . 58, 131–145.
Pubmed Реферат | Pubmed Полный текст
Арредондо, Дж. Т., и Джонсон, Д. А. (2011). Аллометрия ветвления корней и ее связь с морфологическими и функциональными признаками корней у трех сортовых трав. J. Exp. Бот . 62, 5581–5594. DOI: 10.1093 / jxb / err240
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Bengough, A.Дж., Брансби, М. Ф., Ханс, Дж., Маккенна, С. Дж., Робертс, Т. Дж., И Валентин, Т. А. (2006). Реакция корней на физические условия почвы; динамика роста от поля к ячейке. J. Exp. Бот . 57, 437–447. DOI: 10.1093 / jxb / erj003
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Бенго, А. Г., Кастриньяно, А., Пажес, Л., и Ван Нордвейк, М. (2000). «Стратегии выборки, масштабирование и статистика», в Root Methods — A Handbook , под ред. А. Л. Смита, А.Дж. Бенго, К. Энгельс, М. Ван Нордвейк, С. ван де Пеллерин и С. К. Гейн (Берлин: Springer), 147–174.
Pubmed Реферат | Pubmed Полный текст
Bodner, G., Scholl, P., Loiskandl, W., and Kaul, H.-P. (2012). «Можно ли управлять гидравлическими свойствами почвы с помощью корней покровных культур?» В Корни будущего, , изд. Международное общество изучения корней (Данди: 8-й симпозиум Международного общества изучения корней).
Бресинский, А., Кёрнер, Ч., Кадерайт, Дж.W., Neuhaus, G., and Sonnewald, U. (2008). Strasburger — Lehrbuch Der Botanik, 36. Auflage . Гейдельберг: Spektrum Akademischer Verlag.
Кэннон, У.А. (1949). Предварительная классификация корневых систем. Экология 30, 542–548.
Хлоупек, О., Достал, В., Стеда, Т., Псота, В., и Дволякова, А. (2010). Засухоустойчивость сортов ячменя в зависимости от размера корневой системы. Растение породы . 129, 630–636. DOI: 10.1111 / j.1439-0523.2010.01801.x
CrossRef Полный текст
Chloupek, О. (1977). Оценка размера корневой системы растения по его электрической емкости. Почва растений . 48, 525–532. DOI: 10.1007 / BF02187258
CrossRef Полный текст
Корнуэлл, У. К., и Грабб, П. Дж. (2003). Региональные и местные закономерности в разнообразии видов растений с точки зрения наличия ресурсов. Oikos 100, 417–428. DOI: 10.1034 / j.1600-0706.2003.11697.x
CrossRef Полный текст
Дакворт, Дж.К., Кент, М., и Рамзи, П. М. (2000). Функциональные типы растений: альтернатива описанию сообществ тиксономных растений в биогеографии. Прог. Phys. Геогр . 24, 515–542.
Eissenstat, D. M. (1992). Стоимость и преимущества строительства корней малого диаметра. Дж. Завод Нутрь . 15, 763–782. DOI: 10.1080 / 019209364361
CrossRef Полный текст
Монтажник А. Х. (1987). Архитектурный подход к сравнительной экологии корневых систем растений. Новый Фитол . 106, 61–77.
Монтажник, А. Х. (2002). «Характеристики и функции корневой системы», в Корни растений. Скрытая половина, 3-е изд. ., ред. Я. Вайзел, А. Эшел и У. Кафкафи (Нью-Йорк, Нью-Йорк: Марсель Деккер Инк.), 21–50.
Монтажник А. Х., Стикленд Т. Р., Харви М. Л. и Уилсон Г. В. (1991). Архитектурный анализ корневых систем растений 1. Архитектурные корреляты эффективности эксплуатации. Новый Фитол . 118, 375–382. DOI: 10.1111 / j.1469-8137.1991.tb00018.x
CrossRef Полный текст
Гитай Х. и Нобл И. Р. (1997). «Что такое функциональные типы и как их искать?» В Plant Functional Types. Их значение для свойств экосистем и глобальных изменений , ред. Т. М. Смит, Х. Х. Шугарт и Ф. И. Вудворд (Кембридж: Издательство Университета Международной программы по геосфере-биосфере), 3–19.
Хэтчер, Л. (1994). Пошаговый подход к использованию SASR для факторного анализа и моделирования структурных уравнений .Кэри, Северная Каролина: SAS Institute Inc.
Химмельбауэр, М. Л., Лоискандл, В., и Кастанек, Ф. (2004). Оценка длины, среднего диаметра и площади поверхности корней с использованием двух различных систем анализа изображений. Почва растений 260, 111–120. DOI: 10.1023 / B: PLSO.0000030171.28821.55
CrossRef Полный текст
Hochholdinger, F., Woll, K., Sauer, M., and Dembinsky, D. (2004). Генетическое вскрытие корней кукурузы (zea mays) позволяет выявить программы развития, специфичные для каждого типа корня. Ann. Бот . 93, 359–368. DOI: 10.1093 / aob / mch056
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Hund, A., Trachsel, S., and Stamp, P. (2009). Рост пазушных и боковых корней кукурузы: I. Развитие площадки фенотирования. Почва растений 325, 335–349.
Инграм П. А., Чжу Дж., Шарифф А., Дэвис И. В., Бенфей П. Н. и Элич Т. (2012). Высокопроизводительная визуализация и анализ архитектуры корневой системы Brachypodium distachyon в условиях различной доступности питательных веществ. Philos. Пер. R. Soc. Лондон. B Biol. Sci . 367, 1559–1569.
Pubmed Реферат | Pubmed Полный текст
Кайзер, Х. Ф. (1960). Применение электронно-вычислительных машин для факторного анализа. Educ. Psychol. Meas . 20, 141–151. DOI: 10.1177 / 001316446002000116
CrossRef Полный текст
Кашиваги, Дж., Кришнамурти, Л., Упадхьяя, Х. Д., Кришна, Х., Чандра, С., Вадез, В., и др. (2005). Генетическая изменчивость корневых признаков, позволяющих избегать засухи, в коллекции мини-ядра зародышевой плазмы нута ( Cicer arietinum L.). Euphytica 146, 213–222.
Кенрик П. (2002). «Происхождение корней» в корнях растений. Скрытая половина. 3-е изд. , ред. Ю. Вайсель, А. Эшел и У. Кафки (Нью-Йорк, штат Нью-Йорк: Marcel Dekker Inc), 1–20.
Киндшер К. и Уэллс П. В. (1995). Гильдии растений прерий: многомерный анализ видов прерий на основе экологических и морфологических признаков. Растительность 117, 29–50. DOI: 10.1007 / BF00033257
CrossRef Полный текст
Кучера, Л.(1960). Wurzelatlas Mitteleuropäischer Ackerunkräuter und Kulturpflanzen . Франкфурт-на-Майне: DLG-Verlag.
Кучера, Л., и Лихтенеггер, Э. (1997). Bewurzelung von Pflanzen in Verschiedenen Lebensräumen . Stapfia 49, Гуттенберг; Линц: Land Oberösterreeich.
Кучера, Л., Лихтенеггер, Э., Соботик, М. (2009). Wurzelatlas der Kulturpflanzen Gemäßigter Gebiete mit Arten des Feldgemüsebaus . Франкфурт: DLG Verlag.
Лаворель, С., Макинтайр, С., Ландсберг, Дж., И Форбс, Т. Д. А. (1997). Функциональные классификации растений: от общих групп до конкретных групп на основе реакции на нарушение. Дерево 12, 474–478.
Pubmed Реферат | Pubmed Полный текст
Лейтнер Д., Клепш С., Боднер Г. и Шнепф А. (2010a). Модель динамического роста корневой системы, основанная на тропизмах L-систем и связи с поглощением питательных веществ из почвы. Почва растений 332, 177–192.
Лейтнер, Д., Клепш, С., Книсс, А., и Шнепф, А. (2010b). Алгоритмическая красота корней растений — Lsystemmodel для динамического моделирования роста корней. Math. Комп. Модель. Dyn . 16, 575–587.
Ло Сиу, Г., Ясуи, Ю., Чизмади, И., МакГрегор, С. Э., и Робсон, П. Дж. (2011). Изучение статистических подходов к уменьшению субъективности кластерного анализа с целью определения моделей питания: проект будущего. Am. J. Epidemiol . 173, 956–967. DOI: 10.1093 / AJE / kwq458
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Маншади, А., Хаммер, Г., Кристофер, Дж., И ДеВойл, П. (2007). Генотипические вариации архитектурных характеристик корней проростков и их значение для адаптации к засухе у пшеницы. Почва растений 303, 115–129. DOI: 10.1007 / s11104-007-9492-1
CrossRef Полный текст
Миллиган Дж. У. и Купер М. С. (1985). Изучение процедур определения количества кластеров в наборе данных. Психометрика 50, 159–179. DOI: 10.1007 / BF02294245
CrossRef Полный текст
Наим, С.и Райт, Дж. П. (2003). Выявление влияния биоразнообразия на функционирование экосистем: поиск решений, казалось бы, непреодолимой проблемы. Ecol. Lett . 6, 567–579.
Нагель, К. А., Кастенхольц, Б., Янке, С., ван Дуссхотен, Д., Аах, Т., Мюлих, М., и др. (2009). Температурные реакции корней: влияние на рост, архитектуру корневой системы и влияние на фенотипирование. Функц. Завод Биол . 36, 947–959. DOI: 10.1071 / FP09184
CrossRef Полный текст
Нахфоруш, А., Schuhwerk, D., Bodner, G., Kutschka, S., and Grausgruber, H. (2012). «Корневые характеристики твердой пшеницы и ее родственников», в Труды 62-й конференции , изд. Vereinigung der Pflanzenzüchter und Saatgutkaufleute Österreichs (Gumpenstein: Eigenverlag), 101–103.
Осмонт, К. С., Сибут, Р., Хардтке, К. С. (2007). Скрытые ветви: разработки в архитектуре корневой системы. Ann. Ред. Завод Биол . 58, 93–113. DOI: 10.1146 / annurev.arplant.58.032806.104006
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Паж, Л., Веркамбре, Г., Друэ, Дж. Л., Лекомпт, Ф., Колле, К., и Ле Бот, Дж. (2004). Тип корня: общая модель для изображения и анализа архитектуры корневой системы. Растительная почва 258, 103–119. DOI: 10.1023 / B: PLSO.0000016540.47134.03
CrossRef Полный текст
Пастернак, Т., Рудас, В., Поттерс, Г., и Янсен, М.А.К. (2005). Морфогенные эффекты абиотического стресса: переориентация роста проростков Arabidopsis thaliana . Environ. Exp. Бот . 53, 299–314. DOI: 10.1016 / j.envexpbot.2004.04.009
CrossRef Полный текст
Пьефо, Х. П., Бюкс, А., и Рихтер, К. (2004). Смешанный подход к моделированию рандомизированных экспериментов с повторными измерениями. J. Agron. Обрезать. Sci . 190, 230–247. DOI: 10.1111 / j.1439-037X.2004.00097.x
CrossRef Полный текст
Прусинкевич П. и Линденмайер А. (1990). Алгоритмическая красота растений , Берлин: Springer.
Райх, П. Б. (1993). Согласование очевидных расхождений между исследованиями, относящимися к продолжительности жизни, структуре и функциям листьев в разных формах растений и климате: «Слепой и слон пересказали». Funct. Экол . 7, 721–725. DOI: 10.2307 / 23
CrossRef Полный текст
Шенк, Х. Дж., И Джексон, Р. Б. (2002). Глубина укоренения, распространение боковых корней и подземная / надземная аллометрия растений в экосистемах с ограниченными водными ресурсами. Дж.Экол . 90, 480–494.
Шенк, Х. Дж., И Джексон, Р. Б. (2005). Картирование глобального распространения глубоких корней в зависимости от климата и характеристик почвы. Geoderma 126, 129–140.
Симпсон, М. Г. (2005). Систематика растений . Берлингтон, Массачусетс: Academic Press.
Сперри, Дж. С. (2003). Эволюция водного транспорта и структуры ксилемы. Внутр. Дж. Плант Ски . 164, 115–127. DOI: 10.1086 / 368398
CrossRef Полный текст
Тролль, В.(1943). Vergleichende Morphologie der höheren. Пфланцен . Берлин: Borntraeger
Валладарес Ф., Санчес-Гомес Д. и Завала М. А. (2006). Количественная оценка фенотипической пластичности: преодоление разрыва между эволюционной концепцией и ее экологическими приложениями. Дж. Экол . 94, 1103–1116. DOI: 10.1111 / j.1365-2745.2006.01176.x
CrossRef Полный текст
Вругт, Дж., Хопманс, Дж. У. и Шимунек, Дж. (2001). Калибровка двумерной модели поглощения воды корнями. Почвоведение. Soc. Am. J . 65, 1027–1037. DOI: 10.2136 / sssaj2001.6541027x
CrossRef Полный текст
Вейхер, Э., ван дер Верф, А., Томпсон, К., Родерик, М., Гарнье, Э. и Эрикссон, О. (1999). Вызов Теофраста: общий список основных характеристик растений для функциональной экологии. J. Veg. Sci . 10, 609–620. DOI: 10.2307 / 3237076
CrossRef Полный текст
Вернер Т., Мотыка В., Стрнад М. и Шмюллинг Т. (2001). Регулирование роста растений цитокинином. Proc. Natl. Акад. Sci. США . 98, 10487–10492.
Pubmed Реферат | Pubmed Полный текст
Вестоби М. и Лейшман М. (1997). «Классификация видов растений по функциональным типам», в Функциональные типы растений. Их значение для свойств экосистем и глобальных изменений , ред. Т. М. Смит, Х. Х. Шугарт и Ф. И. Вудворд (Кембридж: серия книг Международной программы геосферы-биосферы, University Press), 104–121.
Чжу Дж., Инграм П. А., Бенфей П. Н., Элич Т. (2011). От лаборатории к полю — новые подходы к фенотипированию архитектуры корневой системы. Curr. Мнение. Завод Биол . 14, 310–317. DOI: 10.1016 / j.pbi.2011.03.020
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст
Зобель, Р. В., и Вайзел, Ю. (2010). Архитектурная таксономия корневой системы растений: основа для корневой номенклатуры. Завод Биосист . 144, 507–512. DOI: 10.1080 / 11263501003764483
CrossRef Полный текст
Приложение
В таблице 1 мы предложили основной набор корневых атрибутов, обеспечивающих исчерпывающее описание корневой системы.На рисунке A1 приведен пример корневых классификаторов на разных иерархических уровнях. Качественные дескрипторы могут быть получены для каждого атрибута, что приведет к составной описательной характеристике данного корневого типа.
Рисунок A1. Примеры возможных описательных категорий для иерархически упорядоченных корневых признаков в полном наборе основных данных, которые будут использоваться для классификации типов корневой системы.
Первоначальное различие проводится на основе аллометрических отношений всего растения, таких как соотношение корень: побеги, которые фиксируют ассимилированное распределение между надземными и подземными органами.Следуя Kutschera и Lichtenegger (1997), основное различие между типами корневой системы ожидается от общей формы корневой системы, которая может быть определена геометрической формой. На геометрическую форму или пространственное распределение влияет тип развития (преобладание первичного корня, количество базальных корней, обширность побеговых корней), а также топологическая связь между осями. Таким образом, эти классификаторы размещаются в следующих иерархических порядках. Морфологическое описание осей с одним корнем через средние дескрипторы или производные атрибуты (например,g., распределение диаметра или частоты) логически следует за последующим уровнем. Наконец, в основу иерархической схемы положены анатомические и физиологические характеристики корня. Пример корневых классификационных атрибутов и отличительных категорий на каждом иерархическом уровне показан в Приложении.
Для стабильного результата классификации важно проверить используемые корневые атрибуты. В рамках представленного нами подхода, основанного на данных, это можно сделать на этапе двумерной проверки.В частности, двухуровневый анализ служит для исследования взаимосвязи между чертами, используемыми в качестве классификаторов. Знание коллинеарных отношений между данными важно для обеспечения согласованности классификации в случае использования разных классификаторов. Это проиллюстрировано на Рисунке A2. Векторы схожего направления выражают совместную изменчивость признаков, в то время как перпендикулярное направление показывает признаки, не связанные друг с другом (рис. A2A). При использовании главных компонентов для кластеризации дендрограммы остаются стабильными до тех пор, пока все основные направления вектора на двумерном графике покрываются признаками.Таким образом, описание общей изменчивости системы сохраняется (рисунок A2B). Таким образом, признаки, показывающие сходное направление вектора, могут быть заменены друг другом.
Рисунок A2. Пример двукратной проверки (A) для обеспечения согласованности последующей группировки с помощью кластерного анализа (B) . Биплоты используются для визуализации взаимосвязи между различными признаками и для определения того, какие признаки важны для сохранения общей вариативности в характеристике многопризнаковой корневой системы (данные эксперимента с видами, см.Измеренная морфология корневой системы; Корень: побег — соотношение сухого вещества корня и побега; соотношение массы, длины и диаметра — индикаторы поперечного распределения, рассчитанные путем деления соответствующего параметра, измеренного в рядах растений и между ними).
Таким же образом можно оценить стабильность по годам или площадкам (повторяемость классификации) (Рисунок A3). Видно, что два признака (корень: побег, наклон распределения по глубине) имеют примерно перпендикулярный угол между своими векторами. Также отличается длина вектора.Таким образом, эти признаки очень чувствительны к условиям окружающей среды (годам, участкам). Диаметр корня и длина корня, напротив, имеют схожие направления вектора и длину, что свидетельствует об их стабильности на протяжении двух лет. Следовательно, можно ожидать, что последние признаки приведут к более высокой повторяемости при использовании для классификации на основе кластеров.
Рисунок A3. Пример двух диаграмм для проверки ожидаемой повторяемости групп при последующей кластеризации.