
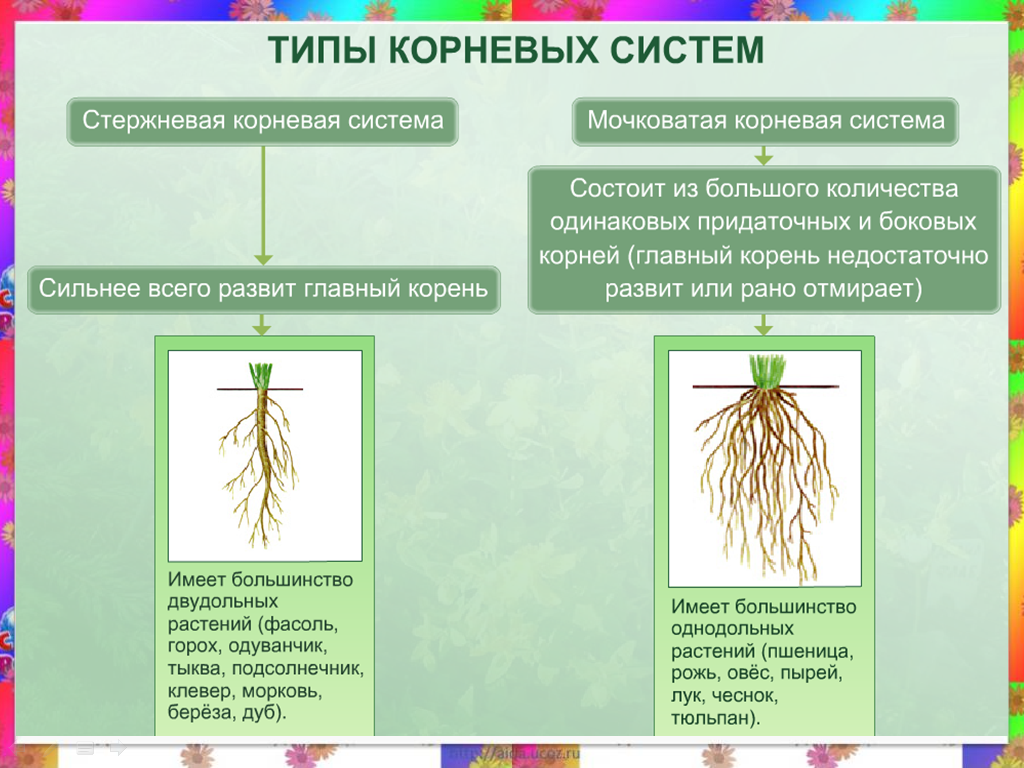
Как получить хорошо развитую корневую систему
Корни ведут скрытую жизнь под землей. На 1 гектаре озимой пшеницы может быть до 300000 км корней, обеспечивающих культуру водой и питательными веществами. Хорошо развитая корневая система — это результат хорошего структурирования почвы, она имеет большое значение для получения высоких урожаев.
Корни удерживают растение в почве и снабжают водой и питательными веществами. Форма и внешний вид корневой системы обычно генетически предопределены, так же как у листьев и стеблей, находящихся на поверхности. Тем не менее, окружающая среда в почве (глина, песок и др.) ограничивает развитие корней. В хорошо-дренированных глинистых почвах, корни растений могут достигать 2-3 м.
Показать больше
Два типа корневых систем
Показать больше
Двудольные, например, масличные культуры, имеют стержневую корневую систему, которая состоит из главного и боковых корней.
Показать больше
Однодольные, например, зерновые, имеют мочковатую, состоящую из 3-5 первичных корней, развивающихся из семени, и придаточных, формирующихся из базальной части стебля. Через 20-30 см неразветвленных корней, растение закладывает очень ветвистую систему.
Показать больше
Высокая скорость, но ограниченная сила
Во время самого активного роста корни двигаются через почву со скоростью примерно 0,5-3 см в день. Однако развитие корней зависит от трещин и пустот, т.к. их способность создавать их собственные каналы ограничена. Во влажной почве вершины корней могут вымещать частицы почвы, но в сухой им необходимо искать каналы с диаметром большим своего. Механическое сопротивление почвы отражается утолщением кончика корня и увеличением разветвленности. Корни и дождевые черви помогают друг другу тем, что корни используют ходы червей, которые в то же время могут воспользоваться старыми корневыми туннелями для движения через почвенный профиль.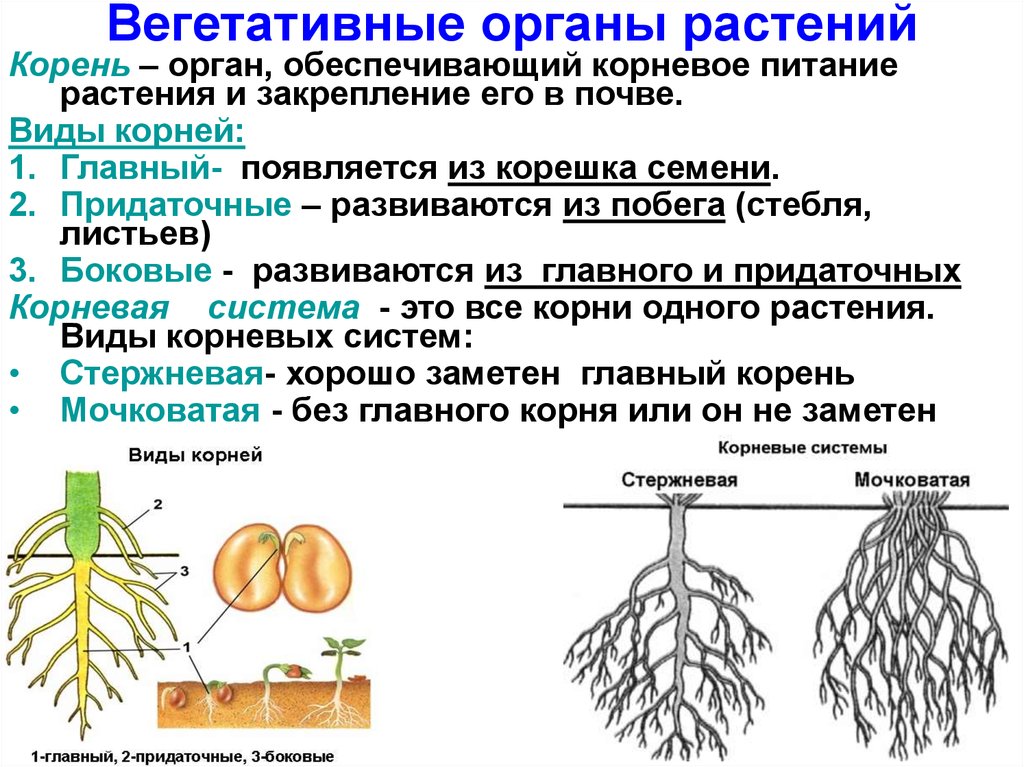
Показать больше
Тонкие нити
Корни очень эффективно всасывают воду и питательные вещества. На кончике корня есть корневой чехлик, и за ним имеется зона деления, где клетки делятся и растут. После зоны деления находится зона всасывания, которая состоит из тоненьких корневых волосков, диаметром около 0,01 мм и длиной 1-10 мм. Эти волоски значительно увеличивают всасывающую способность корней. Всасывающая площадь корней пшеницы, имеющих диаметр около 0,5 мм, за счет корневых волосков может достигать 5 см2 на 1 см длины корня. Вышеописанные волоски покрыты слизью, что еще больше увеличивает контакт с почвой.
Показать больше
100 м корней в одном литре почвы
Эффективность всасывания корневой системой воды и питательных веществ зависит от того, насколько хорошо она проникает через почву. Этот фактор часто зависит от количества длины корней на 1 см3. У зерновых оно равно 10 см/см3, тогда как это число уменьшается до 0,1 см/см3 с увеличением глубины на 1 метр. Значит, верхний слой почвы содержит 100 м корней, когда в одном литре подпочвенного (глубина 1 м от поверхностного) слоя имеется всего 1 м. Длина каждого корня также невероятно велика. Например, под одним м2 поля сахарной свёклы находится около 10 км корней.
Значит, верхний слой почвы содержит 100 м корней, когда в одном литре подпочвенного (глубина 1 м от поверхностного) слоя имеется всего 1 м. Длина каждого корня также невероятно велика. Например, под одним м2 поля сахарной свёклы находится около 10 км корней.
Озимая пшеница имеет еще большую плотность корневой системы: около 30 км корней на м2. Это означает, что один гектар поля озимой пшеницы питается за счет 300 000 км корней, лежащих в почвенном слое.
Показать больше
Словарь:
Однодольные – растения, прорастающие только из одной семядоли (например, спаржа и зерновые).
Двудольные – растения, прорастающие обеими семядолями (например, масличные культуры, горох, бобы, лён, сахарная свёкла).
Подпочва – часть почвенного профиля, находящаяся прямо под поверхностным слоем, и зачастую не зависящая от нормальной обработки почвы (глубокой вспашки), но иногда может затрагиваться глубоким рыхлением.
Показать больше
РОСТ И РАЗВИТИЕ КОРНЕВОЙ СИСТЕМЫ
РОСТ И РАЗВИТИЕ КОРНЕВОЙ СИСТЕМЫ
НА ГЛАВНУЮ / К СОДЕРЖАНИЮ КНИГИ
РОСТ И РАЗВИТИЕ КОРНЕВОЙ СИСТЕМЫ
Рост корневой
система деревьев начинается с момента появления молодых корешков,
независимо от того, каков тип ее происхождения: семенной,
придаточный или отпрысковый. Вначале корни, растут в длину, затем
ветвятся, образуя разные их порядки. У семенных растений рост
корневой системы начинается с развития главного (нулевого порядка)
корня и имеет такую последовательность: от главного корня отходят
корни первого порядка ветвления, от них второго, затем третьего и
последующих порядков ветвления.
 У отпрысковых растений с появлением надземной части корни
уже имеют вполне сформированную систему. У таких растений корневая
система состоит преимущественно из горизонтальных корней и не имеет
стержневого корня, независимо от происхождения корневой системы
маточного растения.
У отпрысковых растений с появлением надземной части корни
уже имеют вполне сформированную систему. У таких растений корневая
система состоит преимущественно из горизонтальных корней и не имеет
стержневого корня, независимо от происхождения корневой системы
маточного растения.Наряду с ростом корней в длину и толщину постоянно происходит их отмирание и возобновление. Явление отмирания 3.А.Колесников (1967) назвал корнепадом. Проведенные им наблюдения на корнях сеянцев яблока и груши показали, что отмирание концов главного и всех длинных боковых корней происходит иногда с первых дней жизни растения. Концы главного корня у 50-85% сеянцев отмирают на 5-20 см длины, после чего образуются боковые корни, которые в процессе роста также подвержены самоизреживанию. Корнепад представляет естественный процесс в жизненном цикле растений. На величину корнепада значительное влияние оказывают, в частности, внешние условия, особенно метеорологические факторы.
С возрастом корневая система увеличивается в диаметре, глубже проникает в почвогрунт, утолщаются скелетные и полускелетные корни, увеличиваются порядки их ветвления, резко возрастает общая протяженность и количество корней.
 Так, по нашим данным, общая
протяженность корней 3-летней яблони сорта Ренет Симиренко, привитой
на лесной яблоне, составила 5285, а 7-летней — уже 12440 м.
Протяженность корней 7-летней яблони сорта Пепинка литовская,
привитой на лесной яблоне и произрастающей в саду по черному пару,
составила (Рубин, 1967) — 14276 м, -14-летней — 26436, 21-летней —
45056 м.
Так, по нашим данным, общая
протяженность корней 3-летней яблони сорта Ренет Симиренко, привитой
на лесной яблоне, составила 5285, а 7-летней — уже 12440 м.
Протяженность корней 7-летней яблони сорта Пепинка литовская,
привитой на лесной яблоне и произрастающей в саду по черному пару,
составила (Рубин, 1967) — 14276 м, -14-летней — 26436, 21-летней —
45056 м.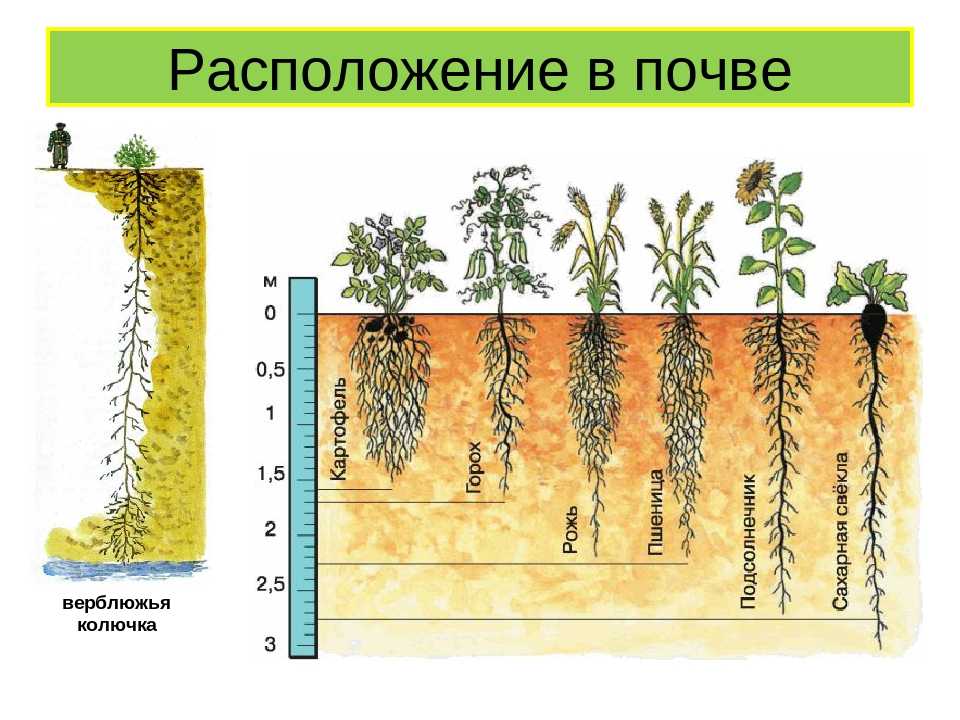 Как видим, на более питательных и лучше увлажненных почвах
доля корней яблони в общей массе растения большая, чем на сухих и
менее питательных почвах.
Как видим, на более питательных и лучше увлажненных почвах
доля корней яблони в общей массе растения большая, чем на сухих и
менее питательных почвах.Таким образом, строение и мощность корневой системы плодовых деревьев обусловливаются как биологическими особенностями подвоя и прививаемого сорта, так и факторами внешней среды. Это требует комплексного изучения корневой системы в конкретных условиях выращивания растений.
НА ГЛАВНУЮ / К СОДЕРЖАНИЮ КНИГИ
Что можно использовать для ускорения роста корней? | Главная Направляющие
Автор: Эшли Маккензи Обновлено 27 декабря 2018 г.
Некоторым растениям нужно что-то, что поможет им укорениться, когда они начинают расти из семян; другим растениям необходимо развить более прочную корневую структуру, которая поддерживает их по мере роста. Хотя вы можете применять сильные стимуляторы корнеобразования в почве вашего сада для быстрого решения проблемы, более разумным вариантом является удобрение с питательными веществами, которые естественным образом стимулируют рост корней.
Хотя вы можете применять сильные стимуляторы корнеобразования в почве вашего сада для быстрого решения проблемы, более разумным вариантом является удобрение с питательными веществами, которые естественным образом стимулируют рост корней.
Питательные вещества для развития корней
Фосфор и калий являются двумя основными питательными веществами, поддерживающими рост корней растений. В частности, они побуждают растения пускать плотный набор новых корней и укреплять существующие корни по мере их развития. Это означает, что удобрения с высоким содержанием фосфора и калия особенно полезны в период активной вегетации. Но прежде чем добавлять в свой сад эти питательные вещества, имейте в виду, что фосфор также стимулирует цветение и производство фруктов — бонус, если вы надеетесь на красочный сад.
Удобрение N-P-K
Коммерческие удобрения обычно имеют соотношение N-P-K, которое указывает процентное содержание азота, фосфора и калия соответственно; если вы хотите удобрение, которое поддерживает рост корней, убедитесь, что второе и третье число больше, чем первое.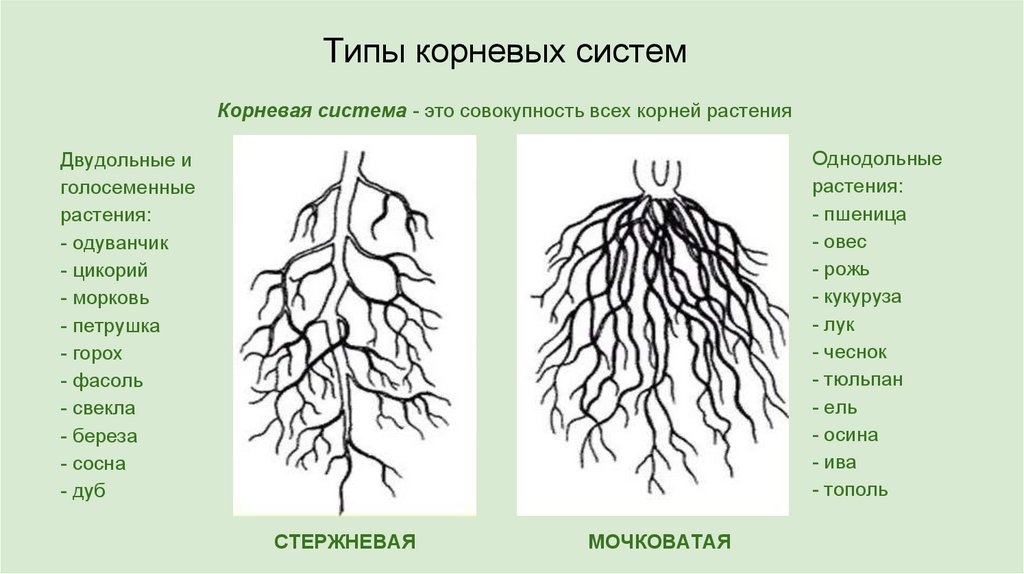 Например, удобрение 3-20-20, содержащее 3 процента азота, 20 процентов фосфора и 20 процентов калия, стимулирует рост корней сильными и здоровыми. Поддерживайте низкое содержание азота, так как он способствует росту длинноногих зеленых растений за счет укоренения, цветения и плодоношения.
Например, удобрение 3-20-20, содержащее 3 процента азота, 20 процентов фосфора и 20 процентов калия, стимулирует рост корней сильными и здоровыми. Поддерживайте низкое содержание азота, так как он способствует росту длинноногих зеленых растений за счет укоренения, цветения и плодоношения.
Природные источники
Природные источники фосфора и калия хорошо подходят для органического сада, и поскольку они обычно высвобождают свои питательные вещества постепенно, они считаются удобрениями с медленным высвобождением. Это означает, что они будут продолжать выделять фосфор и калий для стимулирования развития корневой системы в течение нескольких недель, не вызывая передозировки. Костная мука и каменный фосфат содержат большое количество фосфора и обычно не содержат азота или калия. Между тем, водоросли, гранитная мука, зеленый песок и древесная зола являются отличными источниками калия. Некоторые из этих органических ингредиентов также содержат микроэлементы, такие как железо и магний, полезные для растений.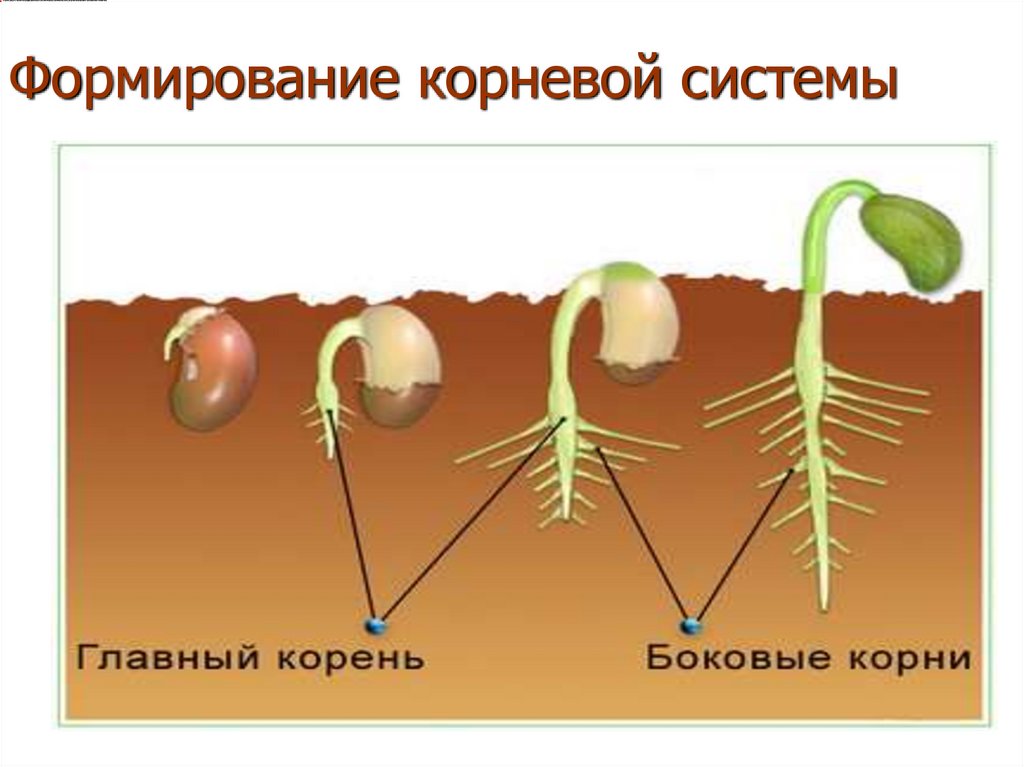
Советы по применению
Если вы хотите стимулировать развитие корней, не вызывая цветения и плодоношения, внесите фосфор и калий перед посадкой. Внесите удобрение на несколько дюймов в почву, где развивающиеся корни смогут его достать. Всякий раз, когда вы применяете фосфор и калий позже, после появления первых ростков, питательные вещества также будут стимулировать рост цветов и плодов.
Стимуляторы корнеобразования
Витамин B1 и растительные гормоны, называемые ауксинами, такие как индолмасляная кислота и нафтилуксусная кислота, считаются стимуляторами корнеобразования. Однако B1 сам по себе не стимулирует рост корней; Единственная причина, по которой некоторые садоводы считают, что это работает, заключается в том, что стимуляторы корнеобразования, содержащие B1, часто также содержат ауксины. Ауксины задерживают рост кроны в пользу развития боковых корней. Это не проблема для укоренившихся растений, но это ядовито для сеянцев, которые нуждаются в меньшем развитии боковых корней, что означает горизонтальное укоренение, которое закрепляет их в почве, и в большем росте первичных корней, что означает глубокие, сильные корни. По этим причинам фосфор и калий обычно являются более безопасными вариантами в саду, особенно для молодых растений.
По этим причинам фосфор и калий обычно являются более безопасными вариантами в саду, особенно для молодых растений.
Ссылки
- Закат: Ускоренный курс по удобрениям
- Arizona Cooperative Extension Backyard Gardener: витамин B-1 и стимуляторы корнеобразования
- Закат: как подкармливать растения
- Урожай к столу: органические удобрения и почва 9003 Поправки 0 Корневое развитие | ПСБ
Корневая система Arabidopsis thaliana – отличная модель для изучения взаимосвязи между регуляцией клеточного цикла, ростом и развитием. Понимание этого открывает большие возможности для изменения архитектуры корней и поглощения воды, что позволяет создавать растения, способные выживать в более засушливых условиях. Исследования этой группы сосредоточены на формировании зачатков боковых корней в перицикле, чтобы выяснить, как регуляция клеточного цикла участвует в инициации новых органов. Корни растений выполняют множество функций.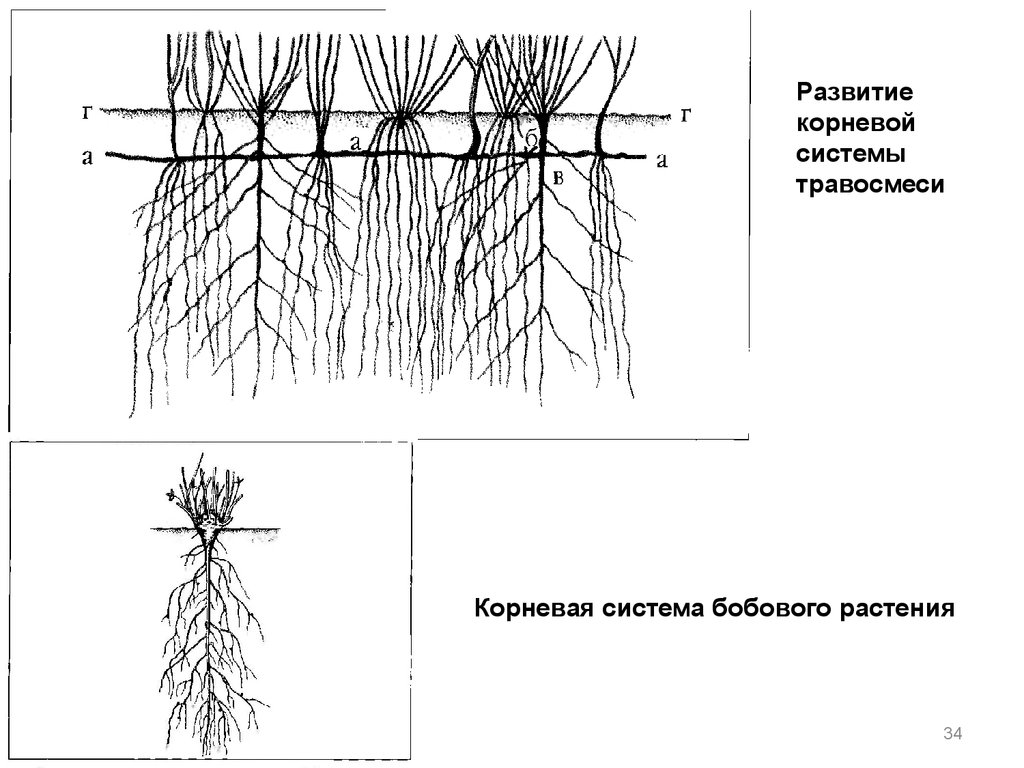
Помимо различных преимуществ этого растения в генетических и молекулярных исследованиях, корень арабидопсиса может похвастаться довольно предсказуемым онтогенезом, очень простой анатомией и высоким уровнем прозрачности, что делает его очень подходящим для морфогенетических и клеточно-биологических исследований. Понимание того, как развиваются корневые системы, имеет решающее значение для максимизации производства сельскохозяйственных культур в мире, в котором растет население, а количество пахотных земель уменьшается.
 Последовательные обработки ингибитором транспорта ауксина и экзогенным ауксином использовались для предотвращения первых формирующих делений и для активации всего перицикла соответственно. Морфологические и молекулярные результаты показывают, что в этой индуцируемой системе активация перицикла соответствует таковой спонтанной инициации латерального корня, но усилена и охватывает всю длину перицикла ксилемы. В этой системе ход клеточного цикла во время ранней индукции боковых корешков отслеживали с использованием гистохимических и молекулярных методов. Результаты показали, что белки, ингибирующие CDK (KRP), играют ранее неизвестную роль в специфическом предотвращении инициации боковых корней при переходе G1-к-S (Himanen et al., в процессе подготовки). Кроме того, анализ паттернов распределения ауксина в системе подтвердил, что ауксин важен для позиционирования и частоты первых формирующих делений для инициации боковых корней (Casimiro et al., 2001).
Последовательные обработки ингибитором транспорта ауксина и экзогенным ауксином использовались для предотвращения первых формирующих делений и для активации всего перицикла соответственно. Морфологические и молекулярные результаты показывают, что в этой индуцируемой системе активация перицикла соответствует таковой спонтанной инициации латерального корня, но усилена и охватывает всю длину перицикла ксилемы. В этой системе ход клеточного цикла во время ранней индукции боковых корешков отслеживали с использованием гистохимических и молекулярных методов. Результаты показали, что белки, ингибирующие CDK (KRP), играют ранее неизвестную роль в специфическом предотвращении инициации боковых корней при переходе G1-к-S (Himanen et al., в процессе подготовки). Кроме того, анализ паттернов распределения ауксина в системе подтвердил, что ауксин важен для позиционирования и частоты первых формирующих делений для инициации боковых корней (Casimiro et al., 2001). Развитие латерального корня (LRD)
У Arabidopsis thaliana закладка латерального корня (LRI) характеризуется скоординированными асимметричными делениями в соседних клетках перицикла полюса ксилемы. Подобно многим процессам развития, он сильно регулируется растительным гормоном ауксином.
Подобно многим процессам развития, он сильно регулируется растительным гормоном ауксином.
Введение
Основным преимуществом перед другими процессами является то, что LRI можно легко манипулировать, что позволило нам разработать систему, индуцируемую латеральным корнем (Himanen et al., 2002). Полногеномный анализ транскрипционных изменений при этой синхронизированной индукции боковых корней привел к выяснению некоторых важнейших аспектов LRI (Himanen et al., 2004 — Cell Cycle; Vanneste et al., 2005 — SLR/IAA14; De Smet et al. ., 2008 – ACR4; De Rybel et al., 2010 – GATA23; De Rybel, Audenaert et al. 2012). Был разработан инструмент, позволяющий объединить эти наборы данных в универсальный сборник (Parizot et al., 2010), что позволило выбрать новые гены-кандидаты для LRI. Эти наборы данных также были доступны в онлайн-инструменте браузера eFP: LRI eFP browser.
Эволюция корней
Изучение развития корней на макро- и микроэволюционном уровне способствовало постоянному совершенствованию методов фенотипирования, расширению баз данных образцов растений и возникновению секвенирования следующего поколения. В группе по развитию корней были инициированы две основные стратегии для исследования микроэволюции архитектуры корней у арабидопсиса и макроэволюции генетических детерминант развития боковых корней у разных видов.
В группе по развитию корней были инициированы две основные стратегии для исследования микроэволюции архитектуры корней у арабидопсиса и макроэволюции генетических детерминант развития боковых корней у разных видов.
Микроэволюция корневой архитектуры арабидопсиса: полногеномное ассоциативное исследование (GWAS)
В последнее время наука о растениях обогатилась разработкой секвенирования следующего поколения, адаптацией статистических инструментов, позволяющих проводить предварительный генетический скрининг растений и дальнейшую эволюцию молекулярных инструменты, доступные для Arabidopsis thaliana. В конечном итоге эти достижения позволяют направить исследования растений на углубленный анализ пула изменчивости генов, который особенно легко доступен и репрезентативен во всем мире для образцов арабидопсиса. На основе данных высокопроизводительного фенотипирования признаков корневой архитектуры in vitro проводятся полногеномные ассоциативные исследования. Это инструмент, используемый для корреляции данных фенотипирования с информацией SNP. Таким образом, мы можем более внимательно изучить, какие гены лежат в основе консервативных подземных фенотипов в выбранных образцах A. thaliana.
Таким образом, мы можем более внимательно изучить, какие гены лежат в основе консервативных подземных фенотипов в выбранных образцах A. thaliana.
Мы дополнительно оцениваем изменчивость и стабильность признаков в наших почвенных, ризотронных системах. Это дает возможность расширить представление об определенных подземных фенотипах и даже связать их с развитием побегов и общей производительностью растения вплоть до стадии цветения.
Макроэволюция развития боковых корней
Большая часть знаний о развитии корней была получена при работе с модельным растением Arabidopsis. Хотя регуляторные механизмы инициации боковых корней и архитектуры корневых систем в значительной степени не раскрыты, неизвестно, насколько это универсально или в какой степени функции генов сохранялись в ходе эволюции. Поэтому, в дополнение к изучению закладки боковых корней у сельскохозяйственных культур, мы изучаем закладку боковых корней и ветвление корней у нескольких эволюционно различных модельных растений.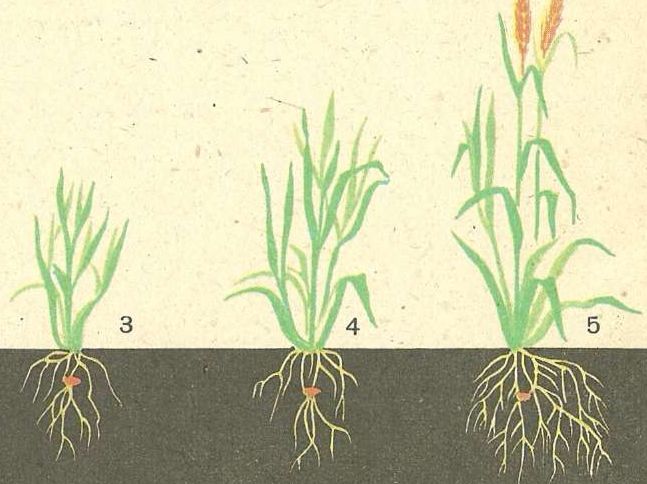 Используемые модели включают раннее наземное растение и ликофит Selaginella moellendorffii, папоротник Ceratopteris richardii (C-папоротник) и покрытосеменные растения, такие как Medicago truncatula и Oryza sativa (рис).
Используемые модели включают раннее наземное растение и ликофит Selaginella moellendorffii, папоротник Ceratopteris richardii (C-папоротник) и покрытосеменные растения, такие как Medicago truncatula и Oryza sativa (рис).
Корни Selaginella произошли независимо от корней высших растений. Они ветвятся не за счет образования боковых корней, а за счет дихотомического сращивания кончика корня. Несмотря на независимую эволюцию, некоторые механизмы развития корней высших растений, по-видимому, переняли и ликофиты. Наше исследование сосредоточено на общепринятых регуляторах как таковых, раскрывающих основные пути развития корней.
У папоротников корень развивается иначе, чем у покрытосеменных. Например, на индукцию боковых корней не влияют ауксины или ингибиторы транспорта ауксинов (как это применяется в системе индуцируемых боковых корней у Arabidopsis).
LRD Transcriptomics
Наборы данных LRI были скомпилированы и интегрированы в онлайн-инструмент браузера eFP. Полногеномный анализ транскрипционных изменений при этой синхронизированной индукции боковых корней привел к выяснению некоторых важнейших аспектов LRI (Himanen et al. , 2004 — Cell Cycle; Vanneste et al., 2005 — SLR/IAA14; De Smet et al. ., 2008 – ACR4; De Rybel et al., 2010 – GATA23; De Rybel, Audenaert et al. 2012). Был разработан инструмент, позволяющий объединить эти наборы данных в универсальный сборник (Parizot et al., 2010), что позволило выбрать новые гены-кандидаты для LRI. Эти наборы данных также были доступны в онлайн-инструменте браузера eFP: LRI eFP browser.
, 2004 — Cell Cycle; Vanneste et al., 2005 — SLR/IAA14; De Smet et al. ., 2008 – ACR4; De Rybel et al., 2010 – GATA23; De Rybel, Audenaert et al. 2012). Был разработан инструмент, позволяющий объединить эти наборы данных в универсальный сборник (Parizot et al., 2010), что позволило выбрать новые гены-кандидаты для LRI. Эти наборы данных также были доступны в онлайн-инструменте браузера eFP: LRI eFP browser.
Сборник наборов данных LRI
Вот снимок относительной экспрессии GATA23 в этих экспериментах (низкие значения экспрессии выделены серым цветом).
LRD в культурных растениях
Среди покрытосеменных существуют значительные различия в строении корня между двудольными и однодольными растениями. Мы работаем над генетическими и молекулярными механизмами, участвующими в развитии корневой системы основных видов сельскохозяйственных культур.
Введение
Из-за различных ролей, которые корневая система играет в общем росте растений, архитектура корня является фундаментальным аспектом роста и развития растений. Корневая система в первую очередь получает воду и питательные вещества из почвы, закрепляет растение в субстрате, синтезирует гормоны и метаболиты, взаимодействует с симбиотическими микроорганизмами и обеспечивает функции запасания. В свете этих характеристик все больше и больше селекционеров сосредотачивают свое внимание на этом подземном органе, чтобы увеличить урожайность. Это требует лучшего понимания связи этой части растения с окружающей средой и ее высоко адаптивного поведения (Lynch, 2007; Gewin, 2010; Den Herder et al., 2010). Среди покрытосеменных существуют большие различия в строении корней между двудольными и однодольными растениями. У двудольных развивается стержневая корневая система, состоящая из основного первичного корня, уже сформировавшегося в эмбриогенезе, который вертикально врастает в почву и дает начало многочисленным боковым корням, расширяющим площадь поверхности. Однодольные имеют мочковатую корневую систему, в которой зародышевый первичный корень важен только для раннего развития растения (Feix et al.
Корневая система в первую очередь получает воду и питательные вещества из почвы, закрепляет растение в субстрате, синтезирует гормоны и метаболиты, взаимодействует с симбиотическими микроорганизмами и обеспечивает функции запасания. В свете этих характеристик все больше и больше селекционеров сосредотачивают свое внимание на этом подземном органе, чтобы увеличить урожайность. Это требует лучшего понимания связи этой части растения с окружающей средой и ее высоко адаптивного поведения (Lynch, 2007; Gewin, 2010; Den Herder et al., 2010). Среди покрытосеменных существуют большие различия в строении корней между двудольными и однодольными растениями. У двудольных развивается стержневая корневая система, состоящая из основного первичного корня, уже сформировавшегося в эмбриогенезе, который вертикально врастает в почву и дает начало многочисленным боковым корням, расширяющим площадь поверхности. Однодольные имеют мочковатую корневую систему, в которой зародышевый первичный корень важен только для раннего развития растения (Feix et al. , 2002), а обширная постэмбриональная корневая система формируется позже. Очень мало известно о генетических и молекулярных механизмах, участвующих в развитии и строении корневой системы основных видов сельскохозяйственных культур, как правило, однодольных растений. Отсутствие понимания, безусловно, является следствием трудности доступа и наблюдения за этим органом в его естественной среде обитания, а именно в почве. Более того, и, вероятно, из-за этого скрытого характера, корень долгое время игнорировался при улучшении сельскохозяйственных культур и в сельскохозяйственных подходах, направленных на увеличение биомассы побегов. Тем не менее, несмотря на то, что большая часть работы была проделана над Arabidopsis thaliana, осознание важности корневой системы в модуляции роста растений вместе с прогрессом в секвенировании и новых молекулярных методах вызвало новый интерес к пониманию молекулярных механизмов у сельскохозяйственных культур ( Hochholdinger and Zimmermann, 2008; Coudert et al.
, 2002), а обширная постэмбриональная корневая система формируется позже. Очень мало известно о генетических и молекулярных механизмах, участвующих в развитии и строении корневой системы основных видов сельскохозяйственных культур, как правило, однодольных растений. Отсутствие понимания, безусловно, является следствием трудности доступа и наблюдения за этим органом в его естественной среде обитания, а именно в почве. Более того, и, вероятно, из-за этого скрытого характера, корень долгое время игнорировался при улучшении сельскохозяйственных культур и в сельскохозяйственных подходах, направленных на увеличение биомассы побегов. Тем не менее, несмотря на то, что большая часть работы была проделана над Arabidopsis thaliana, осознание важности корневой системы в модуляции роста растений вместе с прогрессом в секвенировании и новых молекулярных методах вызвало новый интерес к пониманию молекулярных механизмов у сельскохозяйственных культур ( Hochholdinger and Zimmermann, 2008; Coudert et al.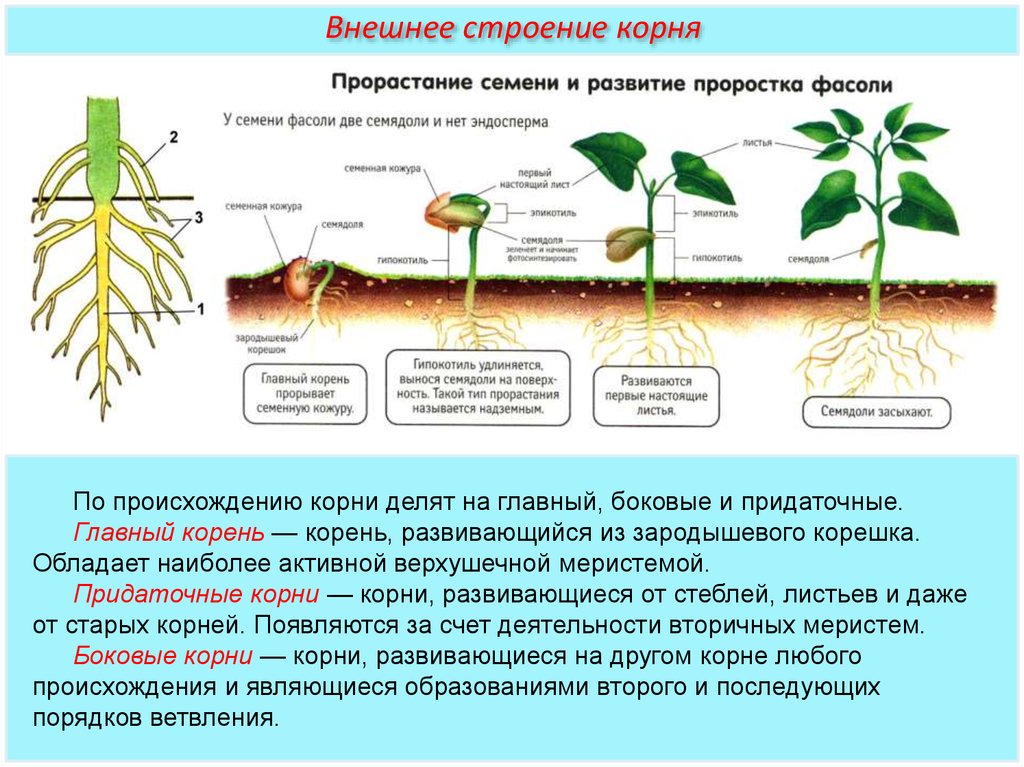 , 2010; Parizot et al., 2012).
, 2010; Parizot et al., 2012).
Ауксин и перицикл
У Arabidopsis thaliana компетентность клеток перицикла в формировании латеральных корней связана с их положением на полюсах ксилемы и зависит от установления максимумов реакции ауксина, локализованных в протоксилеме. У кукурузы наши гистологические анализы выявили прерывание перицикла на полюсах ксилемы и подтвердили ранее сообщавшееся протофлоэмно-специфическое инициирование боковых корней. Клетки перицикла флоэмпола были крупнее и имели более тонкие клеточные стенки по сравнению с другими клетками перицикла, что подчеркивает гетерогенный характер перицикла корня кукурузы. Маркерная линия кукурузы DR5::RFP продемонстрировала наличие максимумов ответа на ауксин в дифференцирующихся клетках ксилемы на кончике корня и в клетках, окружающих сосуды протофлоэмы. Химическое ингибирование транспорта ауксина указывает на то, что установление максимумов реакции на ауксин, локализованных во флоэме, имеет решающее значение для образования боковых корней у кукурузы, поскольку в их отсутствие происходят случайные деления клеток перицикла и эндодермы, не приводящие к органогенезу.