
Строение Корня цветковых растений внешнее, внутреннее и клеточное (Таблица, Схема)
Корень цветковых растений и его строение
| Виды корней | Строение верхушки корня |
|
Внешнее строение |
Внутреннее строение корня |
|
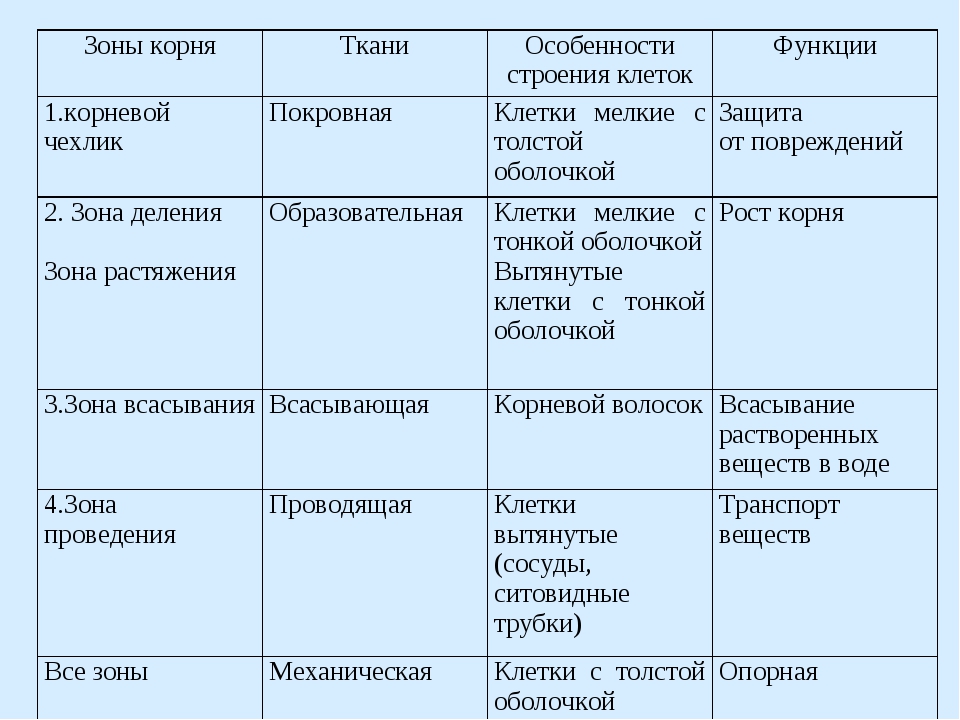
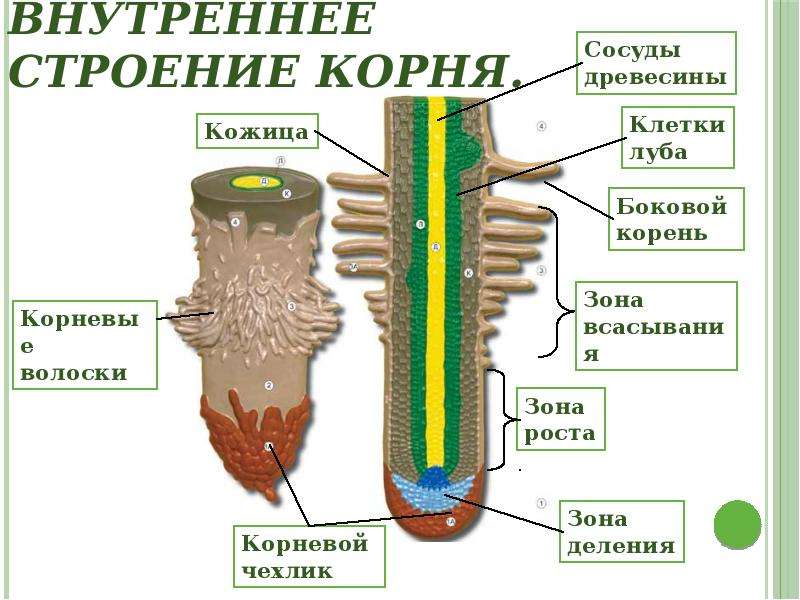
1. Корневой чехлик (мертвые клетки, защищают) 2. Зона деления (мелкие клетки, быстро делятся) 3. Зона роста 4. Зона всасывания (есть выросты — корневые волоски для минерального питания) 5. Зона проведения (проводит минеральные вещества к стеблю) |
1. Кожица (для зашиты») 2. Кора (для зашиты) 3. Камбий (клетки быстро делятся; рост корня) 4. Луб (проводит органические вещества от стебля и листьев) 5. |

Клеточное строение корня
|
Зоны корня |
Клеточное строение корня в зоне всасывания (поперечный срез) |
Схема строения корня в зонах всасывания (1) и проведения (2) |
|
|
|
|
Основные зоны корня |
Описание |
|
зона деления |
Под корневым чехликом располагается зона деления корня. Ее длина около 1 мм. Корень в этой зоне состоит из мелких клеток с тонкими оболочками и крупными ядрами. Они способны делиться в течении всей жизни. |
|
зона растяжения |
В зоне растяжения, или роста, клетки сильно увеличиваются, то есть растягиваются, в продольном направлении (вдоль оси корня). |
|
зона всасывания |
Протяженность зоны всасывания около 2–4 см. На поперечном срезе делится на три участка: кожицу, кору и центральный цилиндр. Кожица корня состоит из тонкостенных клеток. Некоторые из них образуют выросты — корневые волоски, благодаря которым увеличивается всасывающая поверхность корня. Кора корня образована разными клетками. Наружный слой центрального цилиндра состоит из живых тонкостенных клеток, долго сохраняющих способность к делению. Древесина занимает центральную часть корня и ее лучи доходят до внешнего слоя центрального цилиндра. |
|
зона проведения |
Основная функция этой части корня — осуществление нисходящего (по лубу) и восходящего (по древесине) токов растворов веществ. Здесь закладываются и отрастают боковые корни (отсюда второе название — зона ветвления). |
 Их рост происходит главным образом за счет поглощения воды и образования крупных вакуолей. Отдельные клетки этой зоны не только растут, но и делятся.
Их рост происходит главным образом за счет поглощения воды и образования крупных вакуолей. Отдельные клетки этой зоны не только растут, но и делятся.
Основные виды корней
|
Виды корней |
Характеристика |
|
Главный корень |
При про растании семени первым появляется корень. Его называют главным корнем. Он у растения всегда один и формируется из за родышевого корешка. Главный корень быстро растет вертикально вниз. |
|
Боковые корни |
Уже в первую неделю на некотором расстоянии от кончика главного корня образуются боковые корни. |
|
Придаточные корни |
У многих растений, например у пшеницы, ржи, овса, от нижней части стебля отходят мночисленные тонкие корни. Они появляются позднее главного. У некоторых растений (бегонии, сенполии) корни могут образовываться и на листьях. Корни, которые образуются на стебле и листьях, называют придаточными. |
 На придаточных корнях, как и на главном, образуются боковые корни, которые тоже способны ветвиться.
На придаточных корнях, как и на главном, образуются боковые корни, которые тоже способны ветвиться.Типы корневых систем
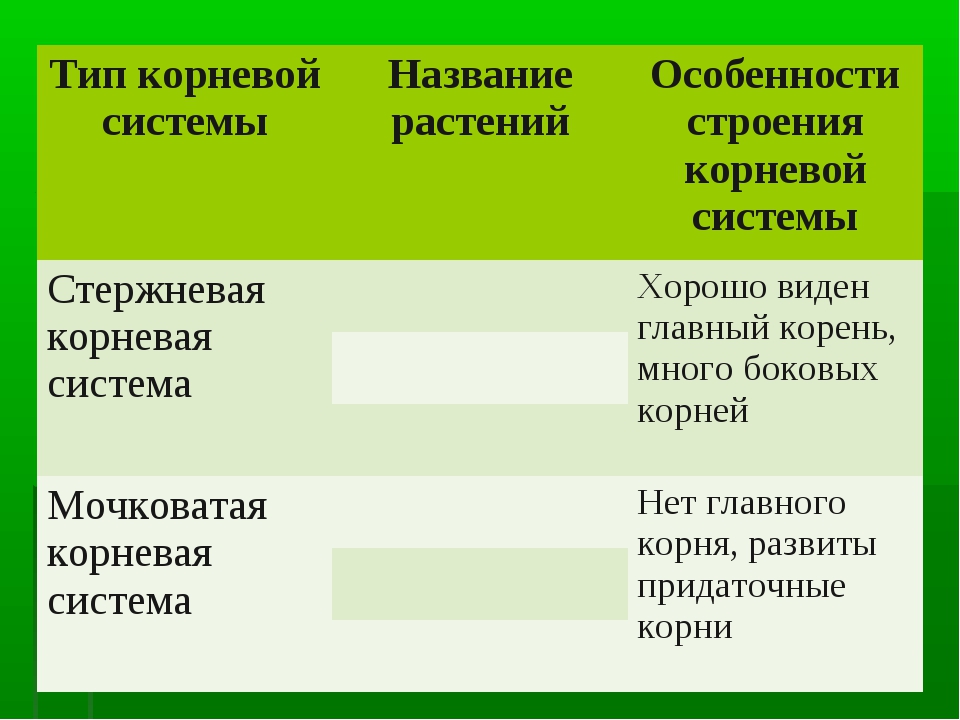
Корневая система — это совокупность всех корней како-голибо растения. Выделяют два основных типа корневых систем, которыеразличаются по форме: стержневая и мочковатая.
|
Типы корневых систем |
Описание |
|
Стержневая |
Хорошо развит главный корень. Он значительно толще и длиннее боковых корней, выглядит как стержень. Распространено у большинства двудольных растений |
|
Мочковатая |
Главный корень не выделяется среди придаточных, развит слабо или вовсе отсутствует. Корни образуют как бы пучок, или мочку. Распространено у однодольных растений |
|
|
Видоизменения корней |
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
Корнеклубни (у георгины) — запас питательных веществ. |
|
Корнеплоды (у моркови, репы) — запас питательных веществ |
|
Воздушные корни (у орхидеи) — для дыхания и питания |
|
Дыхательные корни (у растений топких мест) — для дыхания |
|
_______________
Источник информации:
1. Биология в таблицах и схемах./ Издание 2е, — СПб.: 2004.
2. Биология.Растения.Бактерии.Грибы и лишайники /В.П.Викторов,А.И.Никишов. —М.:ВЛАДОС,2012.—256с.
|
Ткани растений виды |
Строение |
Местонахождения |
Функции |
|
Образовательная (меристема): |
Меристема образована живыми, мелкими, плотными сомкнутыми клетками, с крупным ядром, густой цитоплазмой и мелкими вакуолями |
|
1. 2. Клетки образовательной ткани постоянно делятся и дифференцируются в клетки постоянных тканей. |
|
Первичная верхушечная |
Конус нарастания в почках, зародыше семени, на кончиках корней |
1. Обеспечивает рост органов в длину. 2. Благодаря делению клеток и их дифференциации образуются ткани корней, побегов, листьев, цветков. |
|
|
Вставочная ткань |
Основания междоузлий стебля и основания листа |
||
|
Вторичная боковая (камбий)
|
Расположен между древесиной и лубом стеблей и корней
|
Функция утолщение стебля и корня.
|
|
|
Покровная ткань растений: |
|
Располагается на поверхности |
 Предохраняет растение от высыхания и других неблагоприятных воздействий. 2. Участвует в процессе дыхания. 3. Участвует в обмене веществ между окружающей средой. Предохраняет растение от высыхания и других неблагоприятных воздействий. 2. Участвует в процессе дыхания. 3. Участвует в обмене веществ между окружающей средой.
|
|
Кожица (эпидермис) |
Состоит из слоя живых, плотно сомкнутых клеток с утолщенной стенкой, без хлоропластов. В кожице листьев и зеленых побегов имеются устьица |
Расположена на поверхности листьев, молодых побегов, всех частей цветка |
1. Защита органов от высыхания и микроорганизмов. 2. Устьица обеспечивают газо- и водообмен в растениях. |
|
Пробка (перидерма) |
Состоит из мертвых клеток, стенки которых пропитаны жировым веществом — суберином. Чечевички |
Покрывает зимующие стебли многолетних растений корневища, клубни, корни |
1. Защищает от колебаний температур, механических воздействий, различных вредителей. 2. Многослойная пробка образует на поверхности стебля защитный чехол, в котором находятся чечевички для газо- и водообмена. |
|
Корка (покровный комплекс)
|
Комплекс многослойной пробки и других мертвых тканей, сменяет эпидермис у многолетних растений
|
Покрывает нижнюю часть стволов, хорошо выражена у коркового дуба
|
Выполняет функцию защита от механических повреждений, перепадов температур, вредителей, микроорганизмов.
|
|
Основная ткань — паренхима: |
Основная растительная ткань состоит обычно из живых, тонкостенных клеток, составляющих основу органов |
|
1. Фотосинтез. 2. Запас питательных веществ. 3. Различают также воздухоносную и водоносную паренхимы. |
|
Ассиммиляционая ткань (фотосинтезирующая) |
Столбчатая и губчатая ткань листа, содержит хлоропласты |
В основном — в зеленых листьях и молодых побегах |
1. Фотосинтез 2. Газообмена |
|
Запасащая паренхима |
Состоит из однородных тонкостенных клеток, в которых откладываются белки, жиры, углеводы и другие запасные вещества. Часто имеют крупные вакуоли с клеточным соком |
Она находится в стеблях древесных растений (сердцевина), корнеплодах, клубнях, луковицах, плодах и семенах |
1. Накопление запасных питательных веществ. 2. Клетки основных тканей способны превращаться во вторичную образовательную ткань, за счет которой происходит вегетативное размножение растений. |
|
Водоносная паренхима |
Состоит из крупных, рыхло расположенных клеток |
В стеблях и (или) листьях растений засушливого климата (кактусы, алое, бутылочное дерево) |
Служит для запасания воды у растений засушливого климата |
|
Воздухоносная паренхима — аэренхима
|
Клетки аэренхимы образуют крупные воздухоносные межклетники
|
Развивается у водных и болотных растений в стеблях и реже в листьях (рогоз, тростник)
|
По межклетникам воздух доставляется к подводным частям растений и обеспечивается аэрация
|
|
Проводящая ткань: |
Состоит из вытянутых клеток |
Проводящая ткань является составной частью древесины (ксилемы) и луба (флоэмы) |
Занимается транспортом питательных веществ от корня к листьям (восходящий ток), от листьев к корню (нисходящий) |
|
Ксилема (древесина, сосуды) |
В состав ксилемы входят сосуды (мертвые вытянутые клетки, лишенные поперечных перегородок, стенки которых пропитаны лигнином, придающим сосудам дополнительную твердость), древесинная паренхима и механическая ткань |
Расположена в древесине стебля, проводящей зоне корня, жилках листьев |
Главная проводящая ткань высших сосудистых растений. |
|
Флоэма (луб, ситовидные трубки) |
Состоит из ситовидных трубок с клетками спутниками, лубяной паренхимы и лубяных волокон (механическая ткань). Ситовидные трубки образованы живыми клетками, поперечные перегородки которых пронизаны маленькими отверстиями, образующими «сито». В клетках нет ядер, но они имеют цитоплазму, тяжи которой проходят в соседние клетки через сквозные отверстия в перегородках. Клетки-спутники соединяются с ситовидными трубками плазмодесмами и выполняют функцию питания, синтеза ферментов и так далее. |
Образует проводящие пучки в лубе вдоль стебля, корня, жилок листьев |
Проводит растворенные орган. вещества, образованные в листьях (нисходящий ток), в стебель, корень, цветки, плоды. |
|
Проводящие сосудисто -волокнистые пучки
|
Комплекс из древесины и луба в виде отдельных тяжей у трав и сплошного массива у деревьев
|
Центральный цилиндр корня и стебля; жилки листьев и цветков
|
Проводят по древесине воду и минеральные вещества; по лубу — органические вещества; укрепление органов, связь их в одно целое
|
|
Механическая ткань растений: Волокна |
Клетки механической ткани (лубяные и древесинные волокна) имеют толстые утолщенные и одревесневшие оболочки, плотно прилегающие друг к другу |
Механические ткани в основном расположены в стебле, в корне имеется только в центре. |
Придает прочность органам растения, противодействует разрыву или излому, образуют каркас, поддерживающий органы растения |
|
Каменистые клетки
|
Склереиды — округлые мертвые клетки с очень толстыми одревесневшими оболочками
|
Образуют семенную кожуру, скорлупу ореха
|
Защищают семена от воздействия внешней среды
|
|
Выделительная ткань: |
Состоит из клеток, образующих и выделяющих различные вещества (секреты) |
|
Функция этой ткани растений — выделение секрета |
|
Железистые волоски |
Живые клетки образующие длинные выросты — волоски, внутри которых жидкий секрет |
На поверхности листьев, стеблей (стрекательные клетки крапивы, железистые волоски герани). |
1. Выделение веществ, защищающих от поедания животными, микроорганизмов, испарения 2. Выделение пахучих веществ, привлекающих насе комых- опылителей |
|
Нектарники |
Живые клетки, заполненные сладким веществом, часто сильно пахнущим |
Цветок (чаще всего у основания лепестков) |
Выделение нектара, который привлекает насекомых- опылителей |
|
Смоляные и млечные ходы |
Мертвые вытянуты клетки, заполненные смолой или млечным соком |
Древесина хвойных, стебель одуванчика, молочая |
Защита от микроорганизмов, повреждений, поедания животными |
 Участвует в образовании новых клеток и дифференциации этих клеток в клетки других тканей.
Участвует в образовании новых клеток и дифференциации этих клеток в клетки других тканей.

 Она также участвует в транспорте минеральных веществ (восходящий ток), запасает питательные вещества и выполняет опорную фун-ю
Она также участвует в транспорте минеральных веществ (восходящий ток), запасает питательные вещества и выполняет опорную фун-ю Окружают сосудистые пучки
Окружают сосудистые пучки В основании лепестков
В основании лепестковСтроение Стебля цветковых растений (Таблица)
Основные виды стеблей
|
Вид стебля |
Описание |
|
Прямостоячие |
растут вертикально вверх (береза, подсолнечник) |
|
Вьющиеся |
обвивают опору (фасоль, хмель, вьюнок полевой) |
|
Лазающие |
цепляются за опору усиками или придаточными корнями (горох, плющ) |
|
Ползучие |
стелются по земле и могут укореняться в узлах (земляника) |
Внутреннее строение стебля
|
Однолетний стебель липы срез поперечный |
Строение трехлетней ветки липы |
|
Название зоны стебля |
Тип ткани |
Особенности клеток |
Функции |
|
Кожица |
Покровная |
Один слой клеток, плотно прилегающих друг к другу |
Защита |
|
Чечевички |
Основная |
Крупные клетки с межклетниками |
Газообмен |
|
Пробка |
Покровная |
Мертвые клетки с толстыми стенками |
Защита |
|
Первичная кора |
Покровная |
Клетки плотно прилегают друг к другу |
Защита |
|
Луб |
1. 2. Механическая |
1. Ситовидные трубки 2. Лубяные волокна |
Проведение органических веществ от листьев; опора |
|
Камбий |
Образовательная |
Мелкие клетки, быстро делятся |
Рост стебля в толщину |
|
Древесина |
1. Проводящая 2. Механическая |
1. Сосуды (полые трубки) 2. Древесинные волокна |
Проведение воды и минеральных веществ к листьям; опора. |
|
Сердцевина |
Запасающая |
Крупные клетки с тонкими оболочками и межклетниками |
Запасание веществ |
 Проводящая
Проводящая
|
Передвижение веществ по стеблю |
|
|||
|
|
|
|
|
|
|
|
Минеральных — по сосудам древесины от корня к листьям |
|
Органических — по ситовидным трубкам луба от листьев к органам запасания (корень, луковица) или в центры роста |
|
_______________
Источник информации:
1. Биология в таблицах и схемах./ Издание 2е, — СПб.: 2004.
Биология в таблицах и схемах./ Издание 2е, — СПб.: 2004.
2. Биология.Растения.Бактерии.Грибы и лишайники /В.П.Викторов,А.И.Никишов. —М.:ВЛАДОС,2012.—256с.
Видоизменения корней – таблица типов и строения
Основные функции корня – закрепление в почве растения и поглощение воды. Иногда корни выполняют и другие, не типичные функции. В связи с этим они имеют нетипичное строение, иначе говоря, для таких корней характерны видоизменения, или метаморфозы (от греч. metamorphosis – превращение).
Корнеплоды
Корнеплоды отличаются наличием большого количества запасающей ткани. Они обычно образуются у двулетних растений в первый год жизни. На второй год формируются цветки, плоды, семена. Таким образом, корнеплод позволяет растению перенести период покоя и завершить развитие на следующий год.
Корнеплод – название условное. Он не имеет отношения к плодам, т. к. образуется не из цветка, а из вегетативных органов – стебля и корня.
Соотношение стебля и корня при образовании корнеплодов различно, например, у моркови почти весь корнеплод образован корнем, а у репы стеблем.
Современные корнеплоды выведены человеком искусственно. Они играют важную роль в его питании, а также в кормлении животных.
Корневые шишки
Если корнеплод является утолщённым главным корнем, то корневые шишки – это сильно утолщённые придаточные и боковые корни. Для них, как и для корнеплодов, характерна развитая запасающая паренхима. Корневые шишки образуют придаточные почки, поэтому являются органами вегетативного размножения.
Воздушные корни
Воздушные корни встречаются у многих тропических эпифитов (растений, использующих деревья в качестве опоры).
Рис. 1. Воздушные корни.
Такие корни свободно висят в воздухе и поглощают влагу в виде дождя и росы.
Дыхательные корни
Этот тип видоизменённых корней также встречается в тропиках. Он характерен для деревьев, растущих на заболоченных побережьях океанов. Корневая система таких растений сложна и имеет воздухоносную ткань аэренхиму. Через отверстия воздух попадает в аэренхиму и далее проходит в удалённые подводные участки растения.
Рис. 2. Дыхательные корни.
Ходульные корни
Ходульные корни, или корни-подпорки, образуются у растений, растущих на зыбком илистом грунте. Они распределяют массу растения на увеличенную за их счёт площадь опоры.
Столбовидные корни
Особенность столбовидных корней в том, что они закладываются на ветвях. Характерны такие корни побегов для баньяна.
Баньян – это не название растения. Это название особенности роста некоторых фикусов. Можно назвать баньяном любое дерево с объёмной кроной, опирающееся на столбовидные корни.
Рис. 3. Индийский баньян.
Микориза
Микориза является симбиозом корней и грибов. Она представляет собой корни с проникшими в них гифами гриба. Сожительство с грибом имеет ряд полезных для растения последствий:
- увеличение всасывающей активности корневой системы;
- защита от заражения паразитическими грибами и бактериями;
- увеличение поверхности поглощения воды.
Таблица «Видоизменения корней»
| Типы корней | Примеры растений | Функция |
| Корнеплоды | Репа, редис | запасающая |
| Корневые шишки | Георгин, любка | запасающая |
| Дыхательные | Авиценния | Снабжение воздухом подводных участков растения |
| Ходульные | Ризофора | Увеличение площади опоры |
| Столбовидные | Фикусы | Увеличение площади опоры |
| Корни-зацепки | Плющ и др. лазающие растения лазающие растения | Опорная |
| Корни-присоски | Омела и другие паразиты | Всасывание |
Что мы узнали?
Из статьи по биологии (6 класс) мы узнали, что у многих растений корни выполняют, помимо основных, некоторые дополнительные функции, с чем связаны соответствующие видоизменения корней. Хотя эти функции считаются дополнительными и специфическими, они всё равно имеют отношение к опоре и питанию растения. Типы видоизменённых корней весьма разнообразны.
ПредыдущаяБиологияХвощи и плауны – в чем различие, древние предки, значение
СледующаяБиологияВнешнее строение листа (биология, 6 класс) – характеристика в таблице
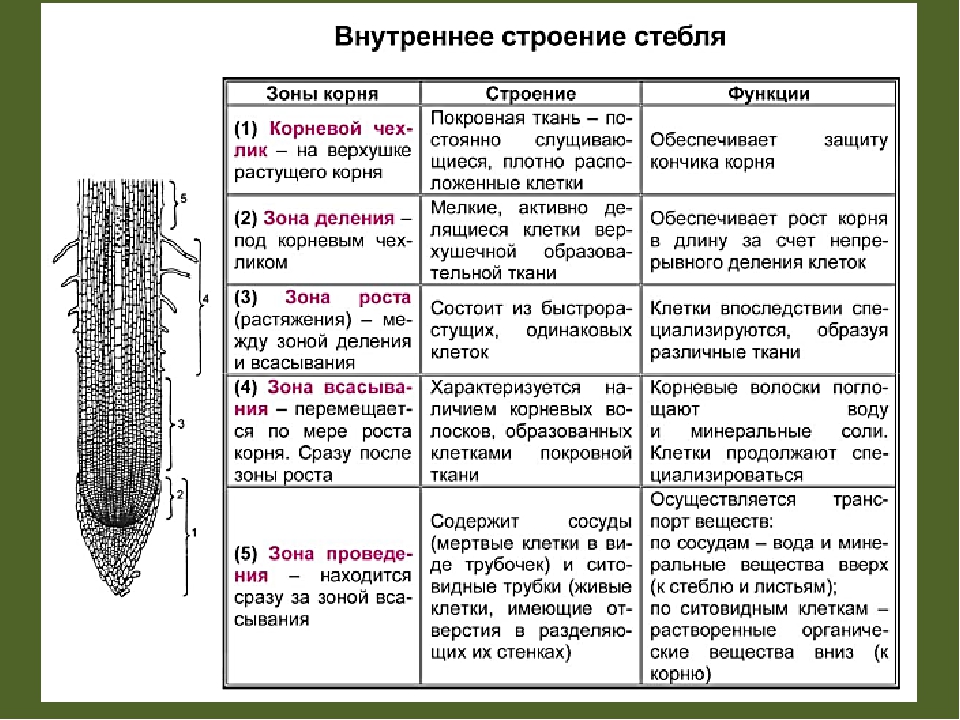
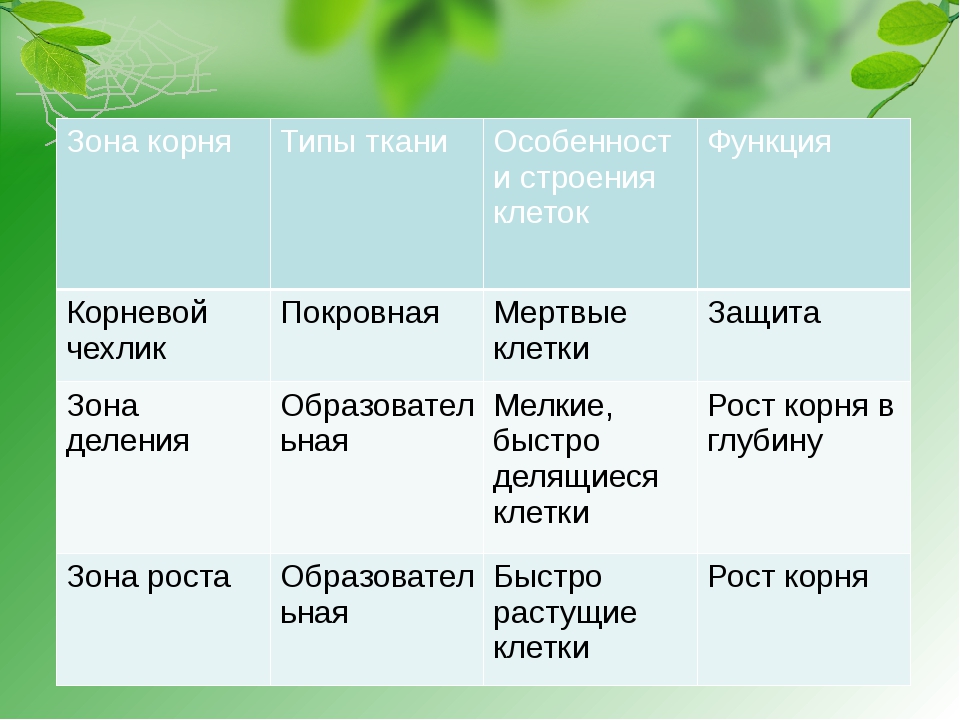
Зоны корня и их функции в таблице
Получение органических веществ и увеличение размера происходит в разных зонах корня. Каждая зона отличается строением, длиной, функцией.
Как расположены зоны
Главный корень развивается из зародыша и растёт исключительно вглубь почвы. Он разделяется на пять зон. Ниже описаны зоны корня по порядку от кончика к стеблю.
Он разделяется на пять зон. Ниже описаны зоны корня по порядку от кончика к стеблю.
Рис. 1. Схема корневых зон.
Место, где корень переходит в стебель, называется корневой шейкой. Обычно это часть тёмная и напоминает по плотности кору.
Корневые зоны и их функции
Клетки каждой зоны отличаются морфологией и функциями. В таблице “Зоны корня и их функции” описаны основные процессы, происходящие в разных частях корня.
Зона | Клетки | Функция |
Чехлик | Уплотнённые, быстро отмирают, выделяют слизь | Защита молодых клеток от повреждений почвы |
Разделения | Мелкие, быстро делящиеся | Происходит непрерывное увеличение корня в длину за счёт деления клеток |
Растяжения | Имеют цилиндрическую, вытянутую форму, мелкие вакуоли сливаются в одну большую | Клетки растягиваются и способствуют продвижению корня вглубь почвы |
Всасывания | Имеют тонкую мембрану и всасывающие волоски | Из почвы с помощью волосков с водой всасываются минеральные вещества |
Боковых корней | Мёртвые и живые, проводящие жидкости | Вверх по стеблям к листьям проводятся питательные вещества, полученные из почвы, а вниз – органические вещества для питания клеток корня. |

Корень растения обладает положительным геотропизмом, т.е. постоянно растёт вниз, к центру Земли. За эту способность отвечают клетки корневого чехлика.
Свойства тканей
Внутреннее строение корня представлено тремя типами ткани:
- эпиблема (ризодерма) – внешняя кожица;
- первичная кора – включает экзодерму, мезодерму, эндодерму;
- центральный, осевой цилиндр или стела – состоит из перицикла и прокамбия.
Рис. 2. Внутреннее строение корня.
Рассмотрим подробнее особенности каждой ткани.
- Эпиблема. Каждый корневой волосок длиной 8-10 мм – это часть клетки ризодермы. Тонкие волоски плотно расположены друг к другу. На один квадратный миллиметр приходится до 300 волосков. Большую часть клетки занимает вакуоль, которую окружает тонкий слой цитоплазмы. Благодаря создающемуся давлению через мембрану в клетку попадает вода и минеральные соли.
 Волоски отмирают через 10-12 дней. За это время в нижней части зоны вырастают новые отростки. Такая смена клеток позволяет волоскам оставаться на одинаковом расстоянии от кончика и постоянно по мере увеличения корня углубляться в почву.
Волоски отмирают через 10-12 дней. За это время в нижней части зоны вырастают новые отростки. Такая смена клеток позволяет волоскам оставаться на одинаковом расстоянии от кончика и постоянно по мере увеличения корня углубляться в почву.
 Волоски отмирают через 10-12 дней. За это время в нижней части зоны вырастают новые отростки. Такая смена клеток позволяет волоскам оставаться на одинаковом расстоянии от кончика и постоянно по мере увеличения корня углубляться в почву.
Волоски отмирают через 10-12 дней. За это время в нижней части зоны вырастают новые отростки. Такая смена клеток позволяет волоскам оставаться на одинаковом расстоянии от кончика и постоянно по мере увеличения корня углубляться в почву.Рис. 3. Строение корневого волоска.
- Первичная кора. Экзодерма содержит более крупные клетки, по сравнению с внутренними слоями. Когда эпиблема отмирает, её место занимает экзодерма. В мезодерме происходит накапливание питательных веществ. Эндодерма образована одним клеточным слоем, опоясывающим осевой цилиндр.
- Стела. Перицикл – верхний слой осевого цилиндра. Прокамбий включает ткани двух типов – ксилему или древесину и флоэму или луб. В состав более плотной ксилемы входят сосуды, доставляющие от волосков питательные вещества в стебли и листья. По тонкому лубу, содержащему ситовидные трубки, приходят органические вещества к каждой клетке корня.
Из-за постоянно растущего кончика клетки постепенно замещают друг друга в разных зонах. Верхние поделившиеся клетки по мере продвижения корня в почву растягиваются и становятся клетками зоны растяжения. Вытянутые клетки всасывают и проводят органические вещества.
Верхние поделившиеся клетки по мере продвижения корня в почву растягиваются и становятся клетками зоны растяжения. Вытянутые клетки всасывают и проводят органические вещества.
Что мы узнали?
Каждый участок корня выполняет определённую функцию за счёт особых клеток, образующих ткани. Зоны позволяют расти внутрь земли, всасывать вещества из почвы и проводить их ко всем остальным растительным частям.
ПредыдущаяБиологияВнешнее строение листа (биология, 6 класс) – характеристика в таблице
СледующаяБиологияКлеточное строение листа – таблица и рисунок для урока в 6 классе
Органы цветковых растений — Сонин 6 класс (ответы)
35. Дайте определение
Орган — это обособленная часть тела, состоящая из тканей и клеток и выполняющая определенные функции
36. Рассмотрите рисунок. Подпишите названия органов цветкового растения, обозначенных цифрами
1. цветок
2.
3. плод
4. лист
5. корень
 стебель
стебель37. Заполните таблицу «Органы цветкового растения»
Название органа Функции Цветок размножение Стебель вынос листоев к свету, опора, транспорт веществ Плод защита и распространение Лист дыхание и фотосинтез, у некоторых: защита и опора Корень укрепление растения в почве, минеральное питание, поглощение воды, запас веществ
38. Выполните задания
1) Корневую систему с хорошо развитым главным корнем называют
стержневой
2) Нарисуйте схему этой системы
3) Корневую систему с недоразвитым или рано отмирающим главным корнем называют
мочковатой
4) Нарисуйте схему этой системы
39. Напишите, у каких растений:
Напишите, у каких растений:
а) стержневая корневая система
у двудольных (горох, дуб, абрикос)
б) мочковатая корневая система
рис, пшеница, лук
40. Рассмотрите рисунок. Подпишите виды корней, обозначенные цифрами
1) Придаточные
2) Боковые
3) Главный
41. Заполните таблицу «Особенности строения корня растения»
Зоны корня Особенности строения Значение для растения Проведения древесина и луб транспорт минеральных веществ из корня в листья и органических веществ обратно Всасывания клетки образуют корневые волоски всасывание минеральных веществ Роста клетки способны делиться образование новых клеток Зона деления клетки способны вытягиваться рост корня Корневой чехлик окружает зону деления защищает зону деления от механического воздействия
42. Рассмотрите рисунок. Назовите видоизменения корней, изображенные на рисунке. Заполните таблицу
Рассмотрите рисунок. Назовите видоизменения корней, изображенные на рисунке. Заполните таблицу
Видоизмененный корень Функции Растения 1. корнеплод запасание питательных веществ редис, морковь, свекла 2. корневый клубни запасание питательных веществ георгины 3. корни-прицепки поднимает растения со слабым стеблем плющ 4. дыхательные корни снабжают кислородом подземные части растения болотный киппарис 5. ходульные корни снабжают кислородом подземные части растения баньян
43. Рассмотрите рисунок. Подпишите части побега, ообзначенные цифрами
1. лист
2. стебель
3. узел
4.
 междоузлие
междоузлие44. Рассмотрите рисунок. Подпиите части стебля, обозначенные цифрами
1. сердцевина
2. древесина
3. камбий
4. кора
5. пробка
45. Заполните таблицу «Строение стебля»
Часть стебля Тип ткани Функции пробка покровная защита, опора луб образовательная транспорт органических веществ камбий проводящая образование новых клеток древесина проводящая транспорт минеральных веществ сердцевина основная запасающая
46. Дайте определение
Почка — это зачаточный побег
47. Зарисуйте листовую и цветочную почки и подпишите их части
Зарисуйте листовую и цветочную почки и подпишите их части
48. Рассмотрите рисунок. Какой лист изображен на рисунке. Подпишите части листа, обозначенные цифрами
1. черешок
2. лист
49. Рассмотрите рисунок. Какими цифрами обозначены простые и сложные листья? Впишите цифры в соответствующие строки
Простые листья: 1, 2, 4, 6, 7, 8,
Сложные листья: 3, 5, 9, 10
50. Дополните предложения
В листе происходят сложные процессы дыхания, образования питательных веществ и испарения воды. У некоторых растений листья выполняют несвойственные им функции: колючки кактуса — защиту, усики гороха — опору
51. Зарисуйте внутреннее строение листа и подпишите его части
52. Рассмотрите рисунок. Подпишите части цветка, обозначенные цифрами
1.
2. Столбик
3. Завязь
4. Цветоножка
5. Цветолистик
6. лепесток
7. Тычиночная нить
8. Пыльник
9. Тычинка
 Рыльце
Рыльце53. Рассмотрите рисунки цветков. Найдите на них цветоножку, цветоложе, чашелистики, лепестки, тычинки, пестики (с завязью, столбиком и рыльцем) и раскрасьте каждую часть цветка одним цветом (например, чашечки зеленым, венчики — желтым, и т.д.)
54. Зарисуйте схему строения обоеполого и раздельнополого цветков
55. Дайте определение
Соцветие — это группа мелких цветков
56. В чем заключается биологическое значение соцветий?
Соцветия необходимы растениям для привлечения насекомых. то повышает вероятность опыления насекомыми
57. Заполните таблицу «Типы соцветий»
58. Заполните схему «Классификация плодов»
Заполните схему «Классификация плодов»
Плоды Сухие Сочные Односемянные Односемянные Примеры:
ясень, одуванчик, подсолнечник
Примеры:
персик, вишня, малина
Многосемянные Многосемянные Примеры:
горох, арахис, дурман
Примеры:
томат, крыжовник, арбуз
59. Рассмотрите рисунок. Подпишите части зерновки кукурузы и семени фасоли, обозначенные цифрами
1. Почечка
2. Стебелек
3. Корешок
4. Семядоли
5. Эндосперм
60. Напишите, какие растения относят к однодольным, а какие — к двудольным
Однодольные: кукуруза, пшеница, лук
Двудольные: капуста, горох, одуванчик
61. Заполните схему «Органы покрытосеменных растений»
Заполните схему «Органы покрытосеменных растений»
ОРГАНЫ вегетативные генеративные корень побег цветок
плод
семя
лист стебель почки
Корни растений: Руководство по основам
Здоровье растения зависит от его корней. Урожайность овощей и фруктов зависит от корнеплодов.
Корни снабжают растение водой и питательными веществами. Корни также поддерживают и прикрепляют растение к почве.
Корень образуется в нижней части зародыша или проростка растения. Кончики корней и крошечные корневые волоски, которые не всегда видны невооруженным глазом, собирают и поглощают пищу и воду из почвы. Более крупные корни, которые легко увидеть, хранят питательные вещества и воду и переносят их в верхние части растения.
Как развиваются корни
Когда прорастающее семя прорастает, из нижней части семени появляется корень и начинает расти прямо вниз. Этот начальный корень называется главным корнем или первичным корнем. Первичный корень может прорасти в почву на несколько дюймов или больше даже до того, как развивающийся побег растения появится на поверхности.
Главный корень может оставаться доминирующим корнем на протяжении всей жизни растения; это касается моркови, шпината и одуванчиков. Или у некоторых растений стержневой корень может быть заменен волокнистыми корнями — массой тонких корней, похожих на веревку; это случай с кукурузой и луком.
У некоторых растений развиваются боковые или боковые корни — корни, которые отходят от исходного главного корня; так обстоит дело с помидорами.
Позже, по мере созревания некоторых растений, у них появляются так называемые придаточные корни; это корни, которые растут из стеблей, побегов и даже листьев. На побегах клубники вырастают придаточные корни.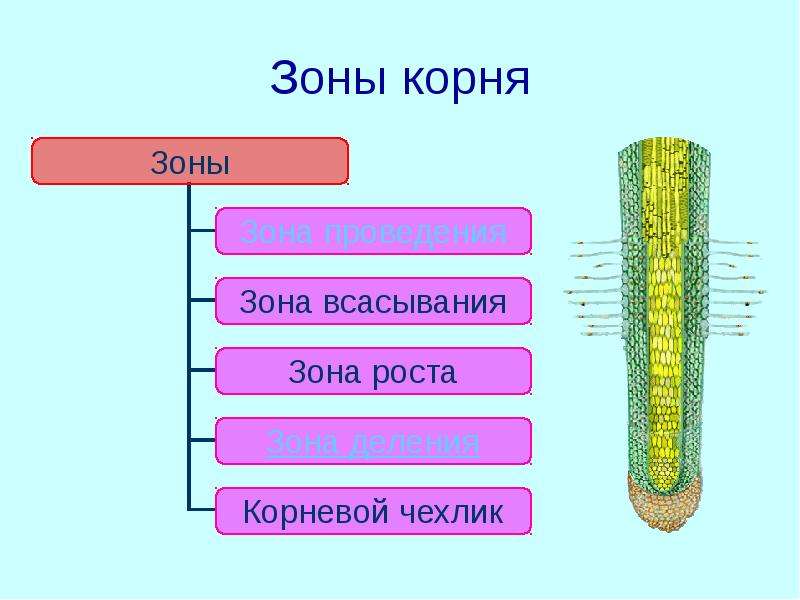
Как укореняется кормовые растения
Корни удерживают растения на месте, прикрепляя их к почве. Но самая важная функция корней — собирать воду и питательные вещества из почвы и помогать растениям расти.
Кончики корней и корневые волоски — самые молодые и самые нежные части корней — постоянно уходят в почву в поисках воды и пищи. Вода и питательные вещества поглощаются кончиками корней и волосками и переносятся к стеблям и листьям через полые клетки, называемые ксилемой. Перекачивание питательных веществ и воды от корней к стеблям и листьям является результатом процесса производства пищи растением, называемого фотосинтезом, который обеспечивается светом.
Растворенные сахара, включая сахар глюкозу, который является основным строительным блоком растительных клеток, и другие материалы, образующиеся в листьях и побегах, циркулируют обратно к корням и через вторую систему полых клеток, называемую флоэмой.Сахар в сочетании с азотом и другими питательными веществами обеспечивает энергию, необходимую корням для роста и функционирования.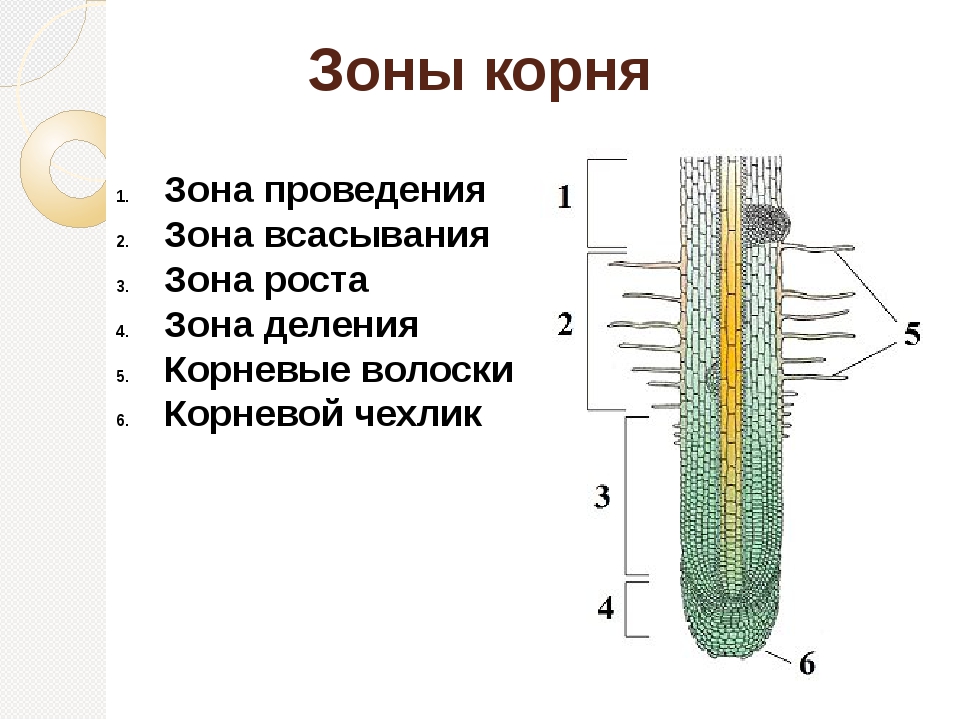
Как почвенные грибы и бактерии взаимодействуют с корнями
Корни некоторых овощей и других растений могут иметь полезные или симбиотические отношения с некоторыми почвенными грибами и бактериями.
Микориза — это структура, которая объединяет мицелий гриба и корень растения. Грибок поглощает растворенные минералы, особенно фосфор, из почвы и передает их корням растений; корень растения, в свою очередь, передает грибу сахара и другие органические соединения, производимые растением.Это симбиотические отношения, известные как микоризная ассоциация.
Определенные виды почвенных бактерий обитают в корнях многих растений семейства бобовых, таких как бобы и клевер. Когда бактерии заражают корень растения, может образоваться круглая опухоль, называемая узелком. Бактерии в клубеньках потребляют часть сахара, производимого растением; в свою очередь, азот, собранный бактериями из почвенного воздуха, превращается в азотные соединения, которые корень растения может поглощать и использовать для питания растения. Этот процесс называется азотфиксацией.Эти бактерии называются азотфиксирующими бактериями.
Этот процесс называется азотфиксацией.Эти бактерии называются азотфиксирующими бактериями.
Статьи по теме:
Питательные вещества для растений
Питательные вещества для овощных растений: источники и дефицит
Щелкните здесь, чтобы получать ежедневные обновления на сайте Harvest to Table в Twitter.
Frontiers | Виртуальным растениям тоже нужна вода: модели функционально-структурной корневой системы в контексте селекции на засухоустойчивость
Введение
Когда в 1960-х годах началась Зеленая революция, население мира составляло 3 миллиарда человек.Примерно 50 лет спустя они достигли 7 миллиардов. Согласно пересмотренным перспективам мирового населения в 2015 году, подготовленным ООН, по медианным оценкам к 2050 году население мира составит почти 10 миллиардов (esa.un.org; Lee, 2011). Прокормить нынешний и будущий мир — это ключевая задача, которая в значительной степени обусловлена возможностью распространения практики Зеленой революции в регионах развивающегося мира, таких как страны Африки к югу от Сахары. Сельскохозяйственное производство во всем мире сталкивается с растущими многофакторными проблемами, среди которых растущее давление на пахотные земли, ухудшение качества почвы, рост цен на удобрения и энергию, а также изменение климата.Что касается только этого последнего пункта, ожидается, что изменения в метеорологической картине (осадки и температура) приведут к снижению средней урожайности всех культур (Knox et al., 2012), особенно из-за засухи, одного из основных ограничений для урожайности сельскохозяйственных культур. (Линч и др., 2014). Это, в свою очередь, отрицательно скажется на продовольственной безопасности в регионах, где основная часть населения борется с хроническим голодом и недоеданием (Schmidhuber and Tubiello, 2007; Lobell et al., 2008; Lobell and Gourdji, 2012).
Сельскохозяйственное производство во всем мире сталкивается с растущими многофакторными проблемами, среди которых растущее давление на пахотные земли, ухудшение качества почвы, рост цен на удобрения и энергию, а также изменение климата.Что касается только этого последнего пункта, ожидается, что изменения в метеорологической картине (осадки и температура) приведут к снижению средней урожайности всех культур (Knox et al., 2012), особенно из-за засухи, одного из основных ограничений для урожайности сельскохозяйственных культур. (Линч и др., 2014). Это, в свою очередь, отрицательно скажется на продовольственной безопасности в регионах, где основная часть населения борется с хроническим голодом и недоеданием (Schmidhuber and Tubiello, 2007; Lobell et al., 2008; Lobell and Gourdji, 2012).
Один из способов борьбы селекционеров с возникающими эпизодами засухи — создание сортов с повышенной устойчивостью к засухе. Это особенно важно для натуральных культур, используемых в развивающихся странах, где люди почти полностью полагаются на эффективную адаптивную способность сельскохозяйственных культур для своего пропитания (Sultan et al.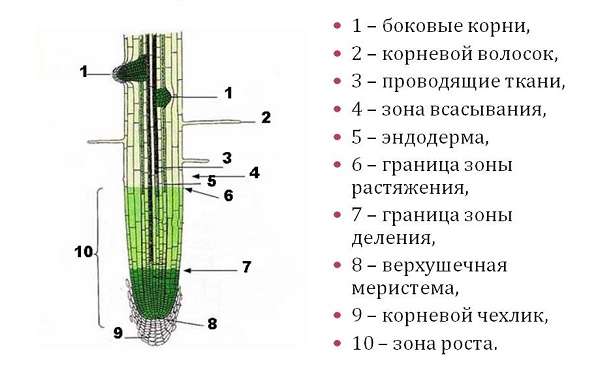 , 2013). Особый интерес в этом контексте представляет корневая система растений. Корни — это органы, отвечающие за поглощение питательных веществ и воды из почвы.Их эффективность зависит от нескольких факторов, главным из которых является их пространственная организация в почве или архитектура корневой системы (RSA) (Den Herder et al., 2010; Draye et al., 2010). RSA является результатом взаимодействия между генетическим программированием развивающихся корней и их реакцией на определенную среду роста (Orman-Ligeza et al., 2014). Следовательно, пластичность развития RSA имеет большое значение, поскольку она определяет приспособляемость растений к ограничениям окружающей среды, например, к условиям засухи (Tuberosa et al., 2002а, б; Линч, 2007; Draye et al., 2010; Тардье, 2012; Leitner et al., 2014b). Другой важный аспект корней, который в значительной степени упускается из виду, — это их функциональность и, в частности, гидравлические процессы, которые способствуют переносу воды и могут объяснить ряд критических фенотипов, измеряемых на уровне побегов, для адаптации к засухе (см.
, 2013). Особый интерес в этом контексте представляет корневая система растений. Корни — это органы, отвечающие за поглощение питательных веществ и воды из почвы.Их эффективность зависит от нескольких факторов, главным из которых является их пространственная организация в почве или архитектура корневой системы (RSA) (Den Herder et al., 2010; Draye et al., 2010). RSA является результатом взаимодействия между генетическим программированием развивающихся корней и их реакцией на определенную среду роста (Orman-Ligeza et al., 2014). Следовательно, пластичность развития RSA имеет большое значение, поскольку она определяет приспособляемость растений к ограничениям окружающей среды, например, к условиям засухи (Tuberosa et al., 2002а, б; Линч, 2007; Draye et al., 2010; Тардье, 2012; Leitner et al., 2014b). Другой важный аспект корней, который в значительной степени упускается из виду, — это их функциональность и, в частности, гидравлические процессы, которые способствуют переносу воды и могут объяснить ряд критических фенотипов, измеряемых на уровне побегов, для адаптации к засухе (см.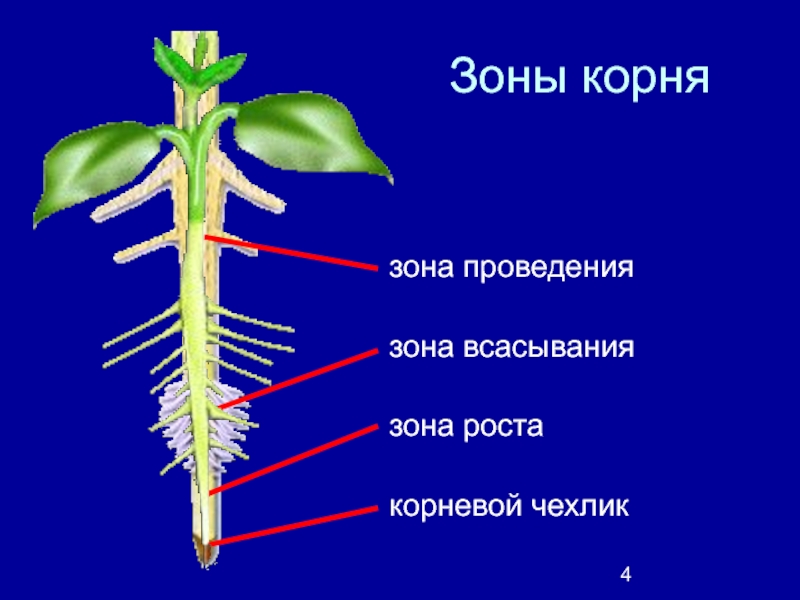 Vadez, 2014).
Vadez, 2014).
Исторические программы селекции были в основном сосредоточены на видимых и легко поддающихся количественной оценке признаках, таких как урожайность зерна, биомасса побегов или устойчивость к болезням и вредителям, все доступные признаки надземных частей растения (Paez-Garcia et al., 2015). Селекционные программы, возникшие в результате Зеленой революции, не были напрямую сосредоточены на корневой системе. Поскольку было показано, что структура корня положительно коррелирует с продуктивностью растений (Lynch, 1995; Kell, 2011; Hufnagel et al., 2014), возрастает интерес к разработке программ селекции растений, направленных на «улучшение» RSA и создание новых сортов с более высокая эффективность использования почвенных ресурсов или лучшая устойчивость к экологическим стрессам, таким как засуха (Wasson et al., 2012).Например, у сорго были идентифицированы QTL для узлового корневого угла, и было показано, что они совмещаются с QTL для признаков, связанных с адаптацией к засухе (Singh et al. , 2010, 2012; Mace et al., 2012). Однако селекция на основе корневых признаков требует выявления специфичных для корней фенов, связанных с оптимизацией исследования почвы и поглощения воды и питательных веществ в различных средах (Lynch et al., 2014). Предпосылкой для этого является возможность фенотипирования и выбора желаемых параметров корня (Paez-Garcia et al., 2015). Однако корневые системы долгое время игнорировались в селекционных программах именно потому, что их сложно фенотипировать. Таким образом, растущий интерес к селекции корней привел к разработке широкого спектра методов фенотипирования корней, охватывающих большую группу условий роста (Paez-Garcia et al., 2015). Эти методы могут использоваться для характеристики и количественной оценки параметров роста и развития корней, необходимых для программ селекции. Примером такой техники в рамках исследования засухи являются лизиметрические системы, в которых растения выращивают в больших трубках, предлагая пространство и объем почвы, аналогичные полевым условиям.
, 2010, 2012; Mace et al., 2012). Однако селекция на основе корневых признаков требует выявления специфичных для корней фенов, связанных с оптимизацией исследования почвы и поглощения воды и питательных веществ в различных средах (Lynch et al., 2014). Предпосылкой для этого является возможность фенотипирования и выбора желаемых параметров корня (Paez-Garcia et al., 2015). Однако корневые системы долгое время игнорировались в селекционных программах именно потому, что их сложно фенотипировать. Таким образом, растущий интерес к селекции корней привел к разработке широкого спектра методов фенотипирования корней, охватывающих большую группу условий роста (Paez-Garcia et al., 2015). Эти методы могут использоваться для характеристики и количественной оценки параметров роста и развития корней, необходимых для программ селекции. Примером такой техники в рамках исследования засухи являются лизиметрические системы, в которых растения выращивают в больших трубках, предлагая пространство и объем почвы, аналогичные полевым условиям. Там корни не извлекаются разрушительным образом, но растительная вода, извлекаемая корнями, то есть их функциональность по отношению к воде, может быть динамически отслежена на протяжении всего цикла урожая (Vadez et al., 2014).
Там корни не извлекаются разрушительным образом, но растительная вода, извлекаемая корнями, то есть их функциональность по отношению к воде, может быть динамически отслежена на протяжении всего цикла урожая (Vadez et al., 2014).
В дополнение к появлению множества разнообразных систем фенотипирования корней, традиционные подходы к селекции теперь могут также извлечь выгоду из понимания механизмов, исходящего из области функционально-структурного моделирования растений (Xu et al., 2011). Подходы к механистическому моделированию дают возможность интегрировать знания о развитии растений и физиологии и оценивать их с учетом разнообразия окружающей среды, что приводит к более надежному селекции (Lynch, 2013, 2015; Lynch et al., 2014). Подходы к функционально-структурной модели растения (FSPM) сосредоточены на моделировании развития, роста и функций всех частей растения (клеток, тканей, органов…) на разном уровне деталей в их пространственно-временном контексте. FSPM — это модели, которые основываются на явном и точном описании рассматриваемой структуры растения, и их эффективность, следовательно, тесно связана с развитием методов фенотипирования. FSPM связывает структуру растения с его физиологическими процессами, которые сами управляются факторами окружающей среды (Godin and Sinoquet, 2005; Vos et al., 2007, 2010). Разработка FSPM носит междисциплинарный характер и использует различные концепции, инструменты и программное обеспечение, происходящие из широкого круга дисциплин (DeJong et al., 2011). В нем могут участвовать ученые с различным опытом: от физиологии растений, развития растений, почвоведения, математики, информатики, клеточной биологии, физики до экологии и агрономии. Например, для кодирования многомасштабной архитектуры растений Годин и Каральо (1998) использовали вложенные графы, которые возникли из математики и широко использовались в других областях, таких как экономика, сети и телекоммуникации, генетика и физика.Помимо кодирования структуры растений, этот многомасштабный формализм также можно использовать для моделирования развития архитектуры растений (Boudon et al., 2012; Ong et al., 2014).
FSPM связывает структуру растения с его физиологическими процессами, которые сами управляются факторами окружающей среды (Godin and Sinoquet, 2005; Vos et al., 2007, 2010). Разработка FSPM носит междисциплинарный характер и использует различные концепции, инструменты и программное обеспечение, происходящие из широкого круга дисциплин (DeJong et al., 2011). В нем могут участвовать ученые с различным опытом: от физиологии растений, развития растений, почвоведения, математики, информатики, клеточной биологии, физики до экологии и агрономии. Например, для кодирования многомасштабной архитектуры растений Годин и Каральо (1998) использовали вложенные графы, которые возникли из математики и широко использовались в других областях, таких как экономика, сети и телекоммуникации, генетика и физика.Помимо кодирования структуры растений, этот многомасштабный формализм также можно использовать для моделирования развития архитектуры растений (Boudon et al., 2012; Ong et al., 2014).
Использование FSPM для моделирования поведения корневой системы сельскохозяйственных культур может помочь понять степень воздействия RSA на данный физиологический процесс (DeJong et al. , 2011). Подходы In silico предлагают преимущество полного владения исследуемой системой и позволяют точно оценить влияние каждого параметра на ее функционирование посредством анализа чувствительности (Han et al., 2012). FSPM, в частности, использовались для моделирования и изучения развития растений в контексте сбора и транспортировки воды (Doussan, 1998; Roose and Fowler, 2004a; Doussan et al., 2006; Javaux et al., 2008; Couvreur et al. , 2012; Lynch et al., 2014). В контексте селекции FSPM могут быть очень полезны, поскольку они используют подход обратного проектирования для определения механизмов растения, которые могут быть полезными в определенных сценариях стрессовой среды. В этом обзоре будут рассмотрены некоторые примеры, показывающие, как имитационные модели сельскохозяйственных культур могут предсказать влияние определенных признаков укоренения на урожайность сельскохозяйственных культур во времени и в географическом масштабе (например,г., Вадез и др., 2013; Холова и др.
, 2011). Подходы In silico предлагают преимущество полного владения исследуемой системой и позволяют точно оценить влияние каждого параметра на ее функционирование посредством анализа чувствительности (Han et al., 2012). FSPM, в частности, использовались для моделирования и изучения развития растений в контексте сбора и транспортировки воды (Doussan, 1998; Roose and Fowler, 2004a; Doussan et al., 2006; Javaux et al., 2008; Couvreur et al. , 2012; Lynch et al., 2014). В контексте селекции FSPM могут быть очень полезны, поскольку они используют подход обратного проектирования для определения механизмов растения, которые могут быть полезными в определенных сценариях стрессовой среды. В этом обзоре будут рассмотрены некоторые примеры, показывающие, как имитационные модели сельскохозяйственных культур могут предсказать влияние определенных признаков укоренения на урожайность сельскохозяйственных культур во времени и в географическом масштабе (например,г., Вадез и др., 2013; Холова и др. , 2014). FSPM также могут служить основой для развития идеотипов, выделяя параметры, которые с наибольшей вероятностью влияют на адаптивность к ограничениям окружающей среды (Lynch, 2013; Lynch et al., 2014).
, 2014). FSPM также могут служить основой для развития идеотипов, выделяя параметры, которые с наибольшей вероятностью влияют на адаптивность к ограничениям окружающей среды (Lynch, 2013; Lynch et al., 2014).
Здесь мы сосредоточены на разработке FSPMs, которые могут использоваться в более широком контексте исследований устойчивости к засухе, связанной с корнями. Сначала мы представим различные методы фенотипирования, существующие для архитектурных и физиологических исследований корней, и их пределы, а также рассмотрим основные черты корня, представляющие интерес для селекционеров.Затем мы представим интеграцию сгенерированных данных в архитектурные модели и то, как эти управляемые данными архитектурные модели могут быть объединены с функциональными гидравлическими моделями, полезными для исследований по разведению. Наконец, мы обсудим оценку и проверку гидравлических моделей FSPM посредством сопоставления моделирования с экспериментами.
Методы фенотипирования корневой системы
Проектирование функционально-структурной модели растения (FSPM) предполагает сбор данных, связанных со структурой растения и физиологическими процессами, которые будут служить исходными данными для питания модели (DeJong et al. , 2011). Фенотипирование растений — это процесс выявления и регистрации качественных и количественных признаков, которые отражают развитие растения и его функциональные аспекты на разных уровнях организации (клетка, ткань, орган, масштаб всего растения) (Granier and Vile, 2014). Стратегии фенотипирования включают навыки и методы, которые позволяют отслеживать развитие растений и их реакцию на различные условия роста, чтобы описать полную архитектурную и / или физиологическую схему во времени и пространстве.За последние годы было разработано множество методов фенотипирования, от лабораторных и тепличных до полевых (Paez-Garcia et al., 2015), и хотя большинство из них были применены к побегам растений (Berger et al., 2012; Araus and Cairns, 2014), некоторые из них позволяют охарактеризовать корневую архитектуру.
, 2011). Фенотипирование растений — это процесс выявления и регистрации качественных и количественных признаков, которые отражают развитие растения и его функциональные аспекты на разных уровнях организации (клетка, ткань, орган, масштаб всего растения) (Granier and Vile, 2014). Стратегии фенотипирования включают навыки и методы, которые позволяют отслеживать развитие растений и их реакцию на различные условия роста, чтобы описать полную архитектурную и / или физиологическую схему во времени и пространстве.За последние годы было разработано множество методов фенотипирования, от лабораторных и тепличных до полевых (Paez-Garcia et al., 2015), и хотя большинство из них были применены к побегам растений (Berger et al., 2012; Araus and Cairns, 2014), некоторые из них позволяют охарактеризовать корневую архитектуру.
Выбор системы фенотипирования корней зависит от нескольких факторов, в том числе от вида растения (однолетнее или многолетнее), целевых представляющих интерес признаков, изученной фазы развития растения (раннее или многолетнее). терминал), необходимость сбора 2D или 3D данных, возможность принести в жертву растение (деструктивные и неразрушающие измерения), временная шкала кинетики роста (дни против месяцев) и затраты (Paez-Garcia et al., 2015) . Разнообразие систем фенотипирования корней, которые разрабатывались годами, теперь позволяет исследователям выбирать установку, наиболее адаптированную к их интересующим вопросам (Kuijken et al., 2015) (Table 1).
терминал), необходимость сбора 2D или 3D данных, возможность принести в жертву растение (деструктивные и неразрушающие измерения), временная шкала кинетики роста (дни против месяцев) и затраты (Paez-Garcia et al., 2015) . Разнообразие систем фенотипирования корней, которые разрабатывались годами, теперь позволяет исследователям выбирать установку, наиболее адаптированную к их интересующим вопросам (Kuijken et al., 2015) (Table 1).
Таблица 1 . Обзор существующих систем фенотипирования корней.
Один простой способ, который может помочь классифицировать и выбирать среди систем корневого фенотипирования, — это рассмотреть их с точки зрения пропускной способности, причем пропускная способность оценивается как масштабированием экспериментальной установки (сколько экспериментальных единиц может быть развернуто параллельно), так и время, необходимое для сбора данных на экспериментальную единицу. Системы фенотипирования на базе лабораторий и теплиц, как правило, позволяют проводить высокопроизводительные эксперименты по фенотипированию (от нескольких сотен до нескольких тысяч растений параллельно и / или быстро собирать данные), что позволяет тестировать большое количество сеянцев в строго контролируемых и воспроизводимых условиях (Таблица 1 ). Эти высокопроизводительные методы имеют решающее значение для исследований QTL или GWAS, направленных на установление связи между пластичностью RSA и генетическими маркерами или конкретными генами или аллелями, которые могут представлять интерес для селекции. Системы со средней пропускной способностью обычно могут работать с десятками растений одновременно и обычно больше фокусируются на пространственном и временном разрешении сбора данных. Независимо от того, являются ли они лабораторными или полевыми, эти системы часто используются для генерации архитектурных и физиологических параметров, используемых как для калибровки, так и для проверки FSPM.На более низком уровне производительности находятся методы, требующие либо дорогостоящих технологических инструментов (например, рентгеновская томография), либо значительного времени сбора данных (например, мелкомасштабная лопатомика). В дополнение к низкой пропускной способности корневая рентгеновская томография до сих пор не освоена полностью, поскольку она подвержена потенциальной потере информации и дополнительному шуму из-за низкого разрешения создаваемых изображений (Mooney et al.
Эти высокопроизводительные методы имеют решающее значение для исследований QTL или GWAS, направленных на установление связи между пластичностью RSA и генетическими маркерами или конкретными генами или аллелями, которые могут представлять интерес для селекции. Системы со средней пропускной способностью обычно могут работать с десятками растений одновременно и обычно больше фокусируются на пространственном и временном разрешении сбора данных. Независимо от того, являются ли они лабораторными или полевыми, эти системы часто используются для генерации архитектурных и физиологических параметров, используемых как для калибровки, так и для проверки FSPM.На более низком уровне производительности находятся методы, требующие либо дорогостоящих технологических инструментов (например, рентгеновская томография), либо значительного времени сбора данных (например, мелкомасштабная лопатомика). В дополнение к низкой пропускной способности корневая рентгеновская томография до сих пор не освоена полностью, поскольку она подвержена потенциальной потере информации и дополнительному шуму из-за низкого разрешения создаваемых изображений (Mooney et al. , 2011) и того факта, что автоматизация Трехмерная реконструкция корневой системы проводится на основе подходов статистического моделирования (Mooney et al., 2011; Kuijken et al., 2015).
, 2011) и того факта, что автоматизация Трехмерная реконструкция корневой системы проводится на основе подходов статистического моделирования (Mooney et al., 2011; Kuijken et al., 2015).
Важным параметром, который следует учитывать при выборе системы фенотипирования корня, является баланс между необходимостью контролируемых условий и наблюдением за «реальным» развитием корня. Лабораторные и тепличные методы, такие как ризотроны, часто ограничивают рост корневой системы до двухмерной структуры ограниченного размера, которая может быстро препятствовать росту корневой системы. Напротив, системы, обеспечивающие постоянный доступ к корню для наблюдения и отбора проб (например,г., гидропоника и аэропоника) означает отсутствие механической среды для поддержки RSA и для воздействия на его развитие, а это означает, что уместность наблюдаемых архитектурных фенотипов в этих установках является дискуссионной. Хотя теоретически менее структурно ограничивающие, полевые методы требуют специальной настройки, такой как укрытия от дождя и ирригационные системы, чтобы обеспечивать контролируемые условия и точно учитывать влияние окружающей среды на развитие корней, а также сильно ограничивать диапазон возможных наблюдений и измерений корневой системы (Паес-Гарсия и др. , 2015). Промежуточные стратегии, такие как ризолизиметры, могут предложить довольно неограниченный рост в контролируемых (или, по крайней мере, контролируемых) условиях, но они требуют значительных структурных инвестиций, чтобы быть практичными.
, 2015). Промежуточные стратегии, такие как ризолизиметры, могут предложить довольно неограниченный рост в контролируемых (или, по крайней мере, контролируемых) условиях, но они требуют значительных структурных инвестиций, чтобы быть практичными.
Процедуры фенотипирования структуры растений обычно можно разделить на три фазы: во-первых, получение архитектурных (и / или физиологических) данных в рамках выбранной системы фенотипирования посредством визуализации, во-вторых, анализ созданного изображения для извлечения количественных данных, касающихся характеров растений. интерес, и, в-третьих, последующий анализ этих количественных данных для извлечения значимой информации, такой как математические законы, описывающие процесс роста.Что касается характеристики корневой архитектуры, первый шаг в основном ограничен трудностью доступа к корневым системам визуально или физически, проблема, для которой было разработано несколько решений (таблица 1). Однако второй шаг, как правило, сильно зависит от возможностей анализа изображений и составляет основное узкое место в исследованиях корневого фенотипирования (Furbank and Tester, 2011).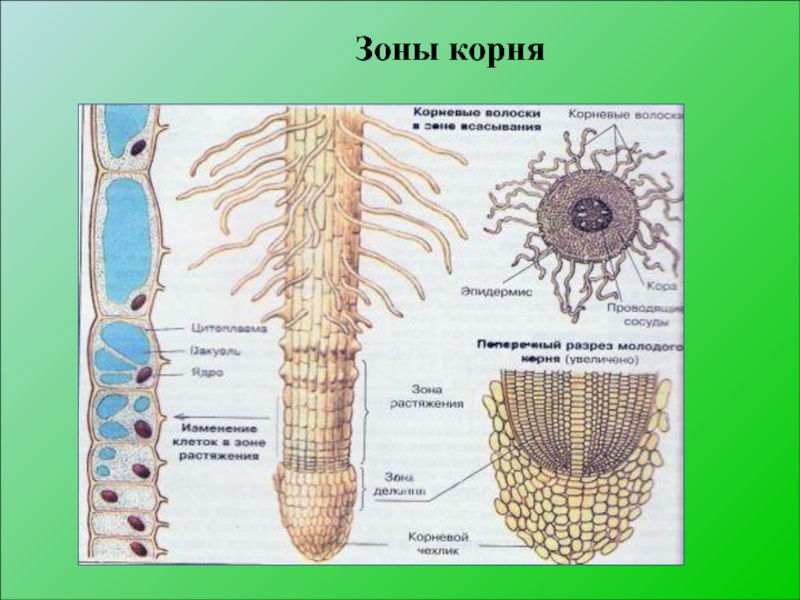
В системах фенотипирования с низкой и высокой пропускной способностью морфологическая и структурная информация в основном генерируется в виде 2D-изображений, которые необходимо преобразовать в количественные данные, представляющие 3D-структуру (Kuijken et al., 2015). А поскольку корневая система представляет собой трехмерную структуру (даже в двухмерном ризотроне), любая достаточно старая корневая система будет демонстрировать перекрытие корней на двухмерных проекционных изображениях. Это значительно усложняет извлечение корневой структуры из изображений и привело к разработке широкого спектра алгоритмов анализа изображений и программных приложений, помогающих автоматически извлекать корневую структуру из зашумленных изображений. Это программное обеспечение для анализа изображений обычно предлагает функции для количественной оценки корневых характеристик, таких как число корней, длина или углы, которые можно использовать для калибровки или проверки корневого FSPM (Godin and Sinoquet, 2005; Vos et al.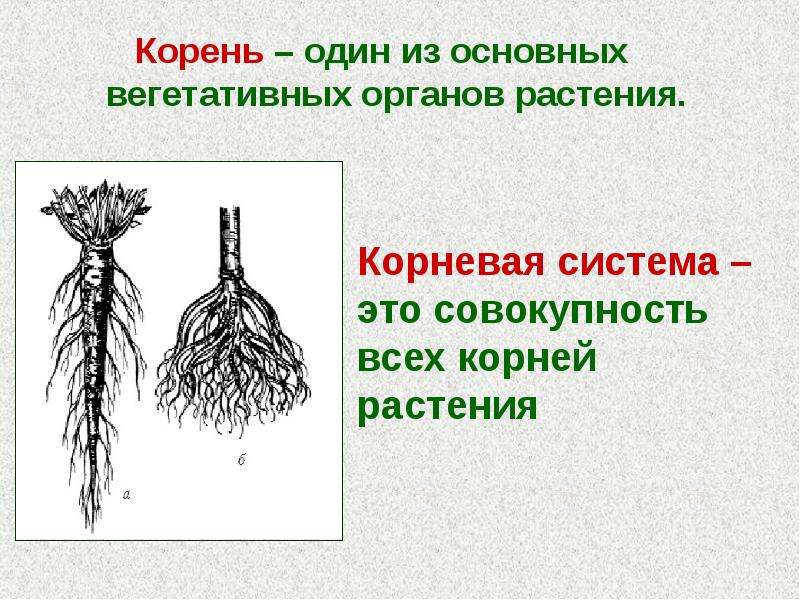 , 2007, 2010; Lynch et al., 2014). Kuijken et al. (2015) недавно сделали обзор всего доступного в настоящее время программного обеспечения для обработки изображений, применимого к корневому фенотипированию. Их увеличивающееся количество приводит к появлению большого разнообразия программных решений для анализа корневых систем (Lobet et al., 2013). Однако это разнообразие также привело к распространению независимых вычислительных методов и фреймворков для представления и хранения корневых архитектур. Это препятствие, ограничивающее возможность обмена данными между лабораториями или использования другого программного обеспечения в одном наборе данных.Чтобы решить эту проблему, недавно было разработано общее корневое описание архитектуры. Появившийся в результате международных совместных усилий нескольких групп, работающих над корневым фенотипированием и моделированием, был определен язык разметки корневой системы (RSML), обеспечивающий переносимость данных корневой архитектуры между программным обеспечением, тем самым способствуя обмену результатами исследований в научном сообществе и создавая стандарт формат, на основе которого строятся центральные хранилища корневых моделей (Lobet et al.
, 2007, 2010; Lynch et al., 2014). Kuijken et al. (2015) недавно сделали обзор всего доступного в настоящее время программного обеспечения для обработки изображений, применимого к корневому фенотипированию. Их увеличивающееся количество приводит к появлению большого разнообразия программных решений для анализа корневых систем (Lobet et al., 2013). Однако это разнообразие также привело к распространению независимых вычислительных методов и фреймворков для представления и хранения корневых архитектур. Это препятствие, ограничивающее возможность обмена данными между лабораториями или использования другого программного обеспечения в одном наборе данных.Чтобы решить эту проблему, недавно было разработано общее корневое описание архитектуры. Появившийся в результате международных совместных усилий нескольких групп, работающих над корневым фенотипированием и моделированием, был определен язык разметки корневой системы (RSML), обеспечивающий переносимость данных корневой архитектуры между программным обеспечением, тем самым способствуя обмену результатами исследований в научном сообществе и создавая стандарт формат, на основе которого строятся центральные хранилища корневых моделей (Lobet et al.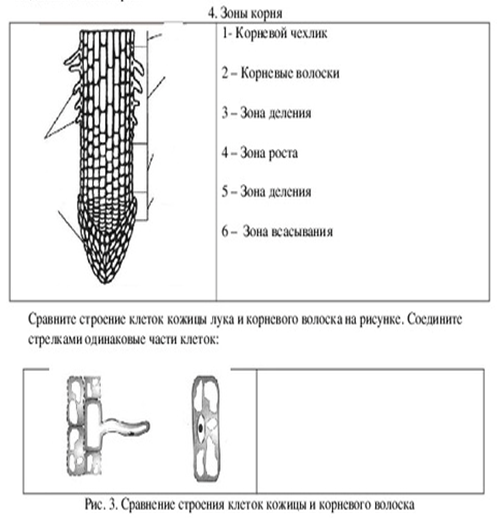 , 2015).
, 2015).
Фенотипирование корневой архитектуры в контексте разведения
Методы фенотипирования, описанные выше, все еще не широко используются в контексте селекционных программ, отчасти потому, что связь между измеряемыми признаками и их полезностью в контексте селекции не всегда очевидна, а отчасти из-за некоторой ограниченной пропускной способности анализа по сравнению к методам анализа геномики, которые могут быть использованы для поддержки программ селекции, таких как, например, GWAS (на основе от тысяч до десятков тысяч растений) (Spindel et al., 2015; Бискарини и др., 2016; Gao et al., 2016; Ивата и др., 2016). Тем не менее, селекционные усилия, направленные на несколько аспектов ЮАР, были успешно предприняты для различных культур (Таблица 2). Например, Tuberosa et al. (2002a, b) идентифицировали QTL для признаков семенных корней в популяции рекомбинантных инбредных линий кукурузы и обнаружили определенную степень совместного расположения между QTL признаков семенных корней и QTL для показателей урожайности при различных водных режимах поля. У нута был идентифицирован основной QTL по признакам корня (глубина, плотность) (Varshney et al., 2014), на основе фенотипических данных, полученных в системе трубок из ПВХ, где растения выращивали, извлекали корни и сканировали через 35 дней после посева (метод см. В Kashiwagi et al., 2005). У сорго была выявлена генотипическая изменчивость узлового корневого угла (Singh et al., 2010, 2012), и эти признаки рассматриваются как потенциальная цель для селекционной программы либо для глубокого укоренения (узкий угол), либо для укоренения в рамках пропущенных — Рядовая посадка, требующая небольшого угла корня. Впоследствии платформа фенотипирования была разработана в масштабе, который позволил фенотипировать картируемую популяцию, и были идентифицированы QTL для узлового корневого угла, и было показано, что они совмещаются с QTL для признаков, связанных с адаптацией к засухе.Эти три примера, взятые из более широкого разнообразия использования корневых признаков в селекции (таблица 2), показывают, как упрощенные методы (например, гидропонная система или измерение угла корня между двумя тонкими пластинами) могут быть достаточными для точного определения генотипической вариации признаков, которые сильно связаны с производительностью на местах.
У нута был идентифицирован основной QTL по признакам корня (глубина, плотность) (Varshney et al., 2014), на основе фенотипических данных, полученных в системе трубок из ПВХ, где растения выращивали, извлекали корни и сканировали через 35 дней после посева (метод см. В Kashiwagi et al., 2005). У сорго была выявлена генотипическая изменчивость узлового корневого угла (Singh et al., 2010, 2012), и эти признаки рассматриваются как потенциальная цель для селекционной программы либо для глубокого укоренения (узкий угол), либо для укоренения в рамках пропущенных — Рядовая посадка, требующая небольшого угла корня. Впоследствии платформа фенотипирования была разработана в масштабе, который позволил фенотипировать картируемую популяцию, и были идентифицированы QTL для узлового корневого угла, и было показано, что они совмещаются с QTL для признаков, связанных с адаптацией к засухе.Эти три примера, взятые из более широкого разнообразия использования корневых признаков в селекции (таблица 2), показывают, как упрощенные методы (например, гидропонная система или измерение угла корня между двумя тонкими пластинами) могут быть достаточными для точного определения генотипической вариации признаков, которые сильно связаны с производительностью на местах.
Таблица 2 . Структурные и функциональные признаки корней определены как потенциально важные для селекции на засухоустойчивость.
Архитектурное моделирование корневой системы
Способность корней обеспечивать гидро-минеральное питание растения зависит от RSA (Lynch, 1995, 2007; Comas et al., 2013; Lynch et al., 2014), но также и от гидравлических характеристик корня. Корневые системы кажутся очень пластичными, а их структура является результатом сложного взаимодействия между генетическими и экологическими регуляторами. Эти взаимодействия порождают динамические петли обратной связи, в которых неоднородность почвенной среды изменяет рост растений, которые, в свою очередь, изменяют почву, собирая из нее воду и питательные вещества, и так далее. Один из способов исследовать и решить такую сложную систему обратной связи — использовать модели.
Поскольку корневые системы спрятаны под землей, моделировать их особенно сложно. Тем не менее, за последние 40 лет было разработано множество архитектурных корневых моделей.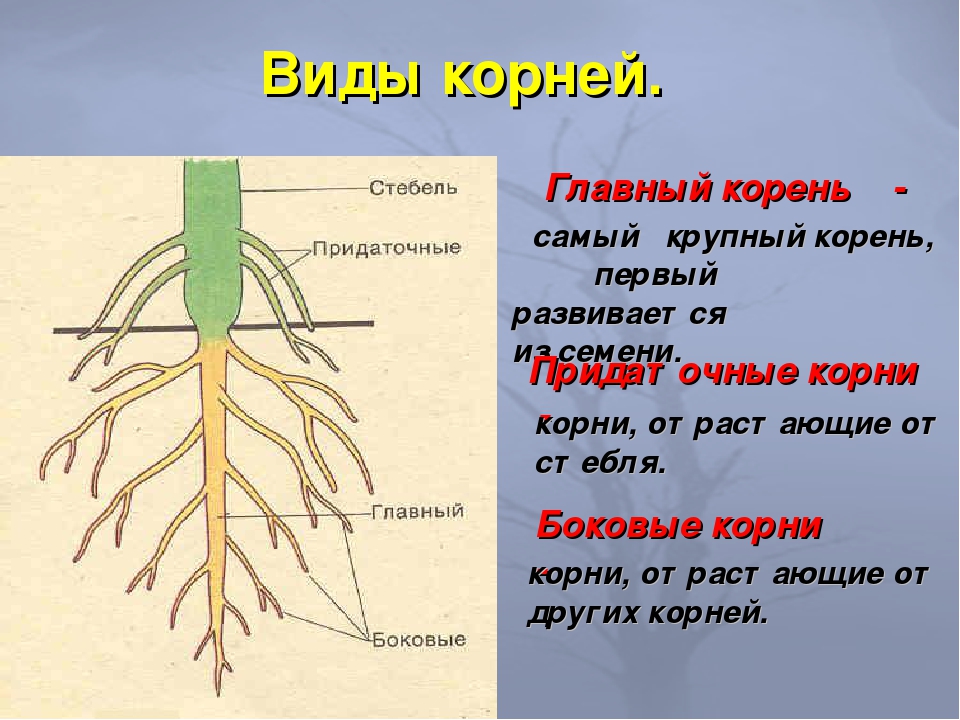 Во всех случаях самый первый шаг процесса моделирования состоит в выборе адекватного представления (то есть формального кодирования) для корневой структуры.
Во всех случаях самый первый шаг процесса моделирования состоит в выборе адекватного представления (то есть формального кодирования) для корневой структуры.
Из-за присущей сложности оценки точного пространственного распределения корней в почве, первая архитектурная модель корневой системы, где фактически непрерывные модели основывались на оценках распределения плотности корней в почве во времени и по глубине (Dupuy et al., 2010). Ранний пример таких моделей использовал уравнение диффузии для моделирования распространения корня в почве (Page and Gerwitz, 1974). Однако модели распределения плотности основывались на синтетических параметрах, таких как единый дескриптор плотности корней, и были основаны на гипотезе о том, что корни равномерно распределяются по почве. Такое предположение не подтверждается в полевых условиях, где наблюдается прерывистое распределение корней, предположительно из-за неоднородного распределения воздействия окружающей среды.Таким образом, простые непрерывные модели не могут легко учесть такие эффекты, как корневая кластеризация, которая играет важную роль в освоении ресурсов (Dupuy et al. , 2010). Как следствие, эти типы непрерывных архитектурных моделей лучше подходят для синтетических описаний RSA в глобальных средах, чем сосредоточение внимания на точном регулировании RSA. Поскольку они могут использоваться для вывода отсутствующей или неточной архитектурной информации, непрерывные модели лучше всего использовать для исследования RSA, когда корневые системы частично или полностью недоступны, например, при изучении взрослых деревьев или растений, выращенных в полевых условиях.Чтобы иметь возможность исследовать RSA в неоднородных почвенных условиях в полевых условиях, непрерывная модель может быть дополнительно дополнена статистическими подходами, позволяющими описывать статистические карты плотности корней в почве (Chopart and Siband, 1999). Основным ограничением этих связанных моделей является их зависимость от данных калибровки, которые должны быть получены из утомительных раскопок in situ и ручного измерения различных частей корневой системы в почве (Chopart and Siband, 1999).
, 2010). Как следствие, эти типы непрерывных архитектурных моделей лучше подходят для синтетических описаний RSA в глобальных средах, чем сосредоточение внимания на точном регулировании RSA. Поскольку они могут использоваться для вывода отсутствующей или неточной архитектурной информации, непрерывные модели лучше всего использовать для исследования RSA, когда корневые системы частично или полностью недоступны, например, при изучении взрослых деревьев или растений, выращенных в полевых условиях.Чтобы иметь возможность исследовать RSA в неоднородных почвенных условиях в полевых условиях, непрерывная модель может быть дополнительно дополнена статистическими подходами, позволяющими описывать статистические карты плотности корней в почве (Chopart and Siband, 1999). Основным ограничением этих связанных моделей является их зависимость от данных калибровки, которые должны быть получены из утомительных раскопок in situ и ручного измерения различных частей корневой системы в почве (Chopart and Siband, 1999).
RSA возникают в результате взаимодействия между корневыми процессами развития и окружающей их средой. Поскольку они не рассматривают отдельные корни, непрерывные модели не могут легко учесть обратную связь, существующую между корнем и почвой. Следовательно, необходимы новые подходы, чтобы понять, как почва исследуется растением на уровне отдельной корневой оси (Pierret et al., 2007). Это соображение привело к разработке более сложных корневых моделей. Эти модели основаны на явном описании процессов развития, роста и ветвления корней, в результате чего получаются одномерные, двухмерные или трехмерные модели (Dunbabin et al., 2013). Такие дискретные и явные модели рассматривают корневую архитектуру через ее полную дискретную топологию и геометрию и могут быть основаны на нескольких различных математических формализмах (Godin, 2000; Balduzzi et al., 2017). Двумя наиболее популярными формализмами, используемыми для представления дискретной архитектуры предприятия в целом, являются многомасштабные древовидные графы (MTG) (Godin and Caraglio, 1998; Godin et al. , 1999, 2005) и L-системы (Prusinkiewicz and Lindenmayer, 1990). .
, 1999, 2005) и L-системы (Prusinkiewicz and Lindenmayer, 1990). .
MTG были разработаны на основе концепции модульности растений и нацелены на описание отдельных частей растений в виде древовидных графов, которые сами включены в древовидную структуру (Godin et al., 1997). MTG допускают топологическое и геометрическое кодирование любого вида растений и использовались в качестве стандарта для описания развития растений и архитектуры широкого круга видов (Godin and Caraglio, 1998; Danj
Frontiers | Корневой экссудат первичных метаболитов: механизмы и их механизмы) Роль в реакции растений на воздействия окружающей среды
- 1 Исследования наземных экосистем, Департамент микробиологии и экосистемных наук, Исследовательская сеть «Химия и микробиология», Венский университет, Вена, Австрия
- 2 Факультет естественных наук, Сиднейский институт сельского хозяйства, Сиднейский университет, Сидней, Новый Южный Уэльс, Австралия
Экссудация корней — важный процесс, определяющий взаимодействие растений с почвенной средой.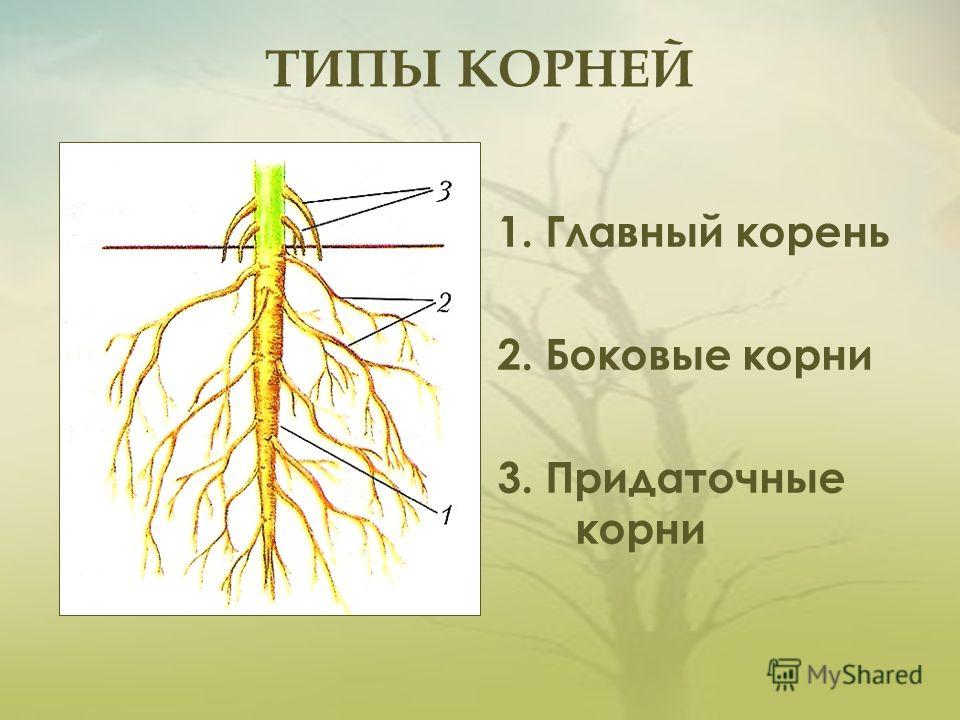 Многие исследования связывают этот процесс с мобилизацией питательных веществ в почве. Тем не менее, остается нерешенным, как контролируется экссудация и как именно и при каких обстоятельствах растения получают пользу от экссудации. Считается, что большинство корневых экссудатов, включая первичные метаболиты (сахара, аминокислоты и органические кислоты), пассивно теряются из корня и используются микробами, обитающими в ризосфере. В этом обзоре мы синтезируем последние достижения в экологии и биологии растений, чтобы объяснить и предложить механизмы, с помощью которых контролируется выделение первичных метаболитов корнями, и какую роль их экссудация играет в стратегиях усвоения питательных веществ растениями.В частности, мы предлагаем новую концептуальную основу для корневых экссудатов. Эта структура построена на двух основных концепциях: (1) корневое выделение первичных метаболитов обусловлено диффузией, при этом растения и микробы модулируют градиенты концентрации и, следовательно, скорость диффузии в почву в зависимости от их статуса питания; (2) концентрация выделяемых метаболитов может быть определена на кончике корня, и сигналы транслируются для изменения структуры корня.
Многие исследования связывают этот процесс с мобилизацией питательных веществ в почве. Тем не менее, остается нерешенным, как контролируется экссудация и как именно и при каких обстоятельствах растения получают пользу от экссудации. Считается, что большинство корневых экссудатов, включая первичные метаболиты (сахара, аминокислоты и органические кислоты), пассивно теряются из корня и используются микробами, обитающими в ризосфере. В этом обзоре мы синтезируем последние достижения в экологии и биологии растений, чтобы объяснить и предложить механизмы, с помощью которых контролируется выделение первичных метаболитов корнями, и какую роль их экссудация играет в стратегиях усвоения питательных веществ растениями.В частности, мы предлагаем новую концептуальную основу для корневых экссудатов. Эта структура построена на двух основных концепциях: (1) корневое выделение первичных метаболитов обусловлено диффузией, при этом растения и микробы модулируют градиенты концентрации и, следовательно, скорость диффузии в почву в зависимости от их статуса питания; (2) концентрация выделяемых метаболитов может быть определена на кончике корня, и сигналы транслируются для изменения структуры корня. Поток первичных метаболитов через корневую экссудацию в основном находится на кончике корня, где отсутствие дифференцировки клеток способствует диффузии метаболитов в почву.Мы показываем примеры того, как кончик корня воспринимает изменения концентрации выделяемых метаболитов и преобразует это в сигналы, чтобы изменить рост корня. Растения могут изменять концентрацию метаболитов либо путем управления процессами источника / поглощения, либо путем экспрессии и регулирования переносчиков оттока, тем самым ставя под сомнение идею корневой экссудации как чисто нерегулируемого пассивного процесса. Посредством оттока корневого экссудата растения могут локально увеличивать концентрацию многих общих метаболитов, которые могут служить датчиками и интеграторами состояния питания растений и доступности питательных веществ в окружающей среде.Связанные с растениями микроорганизмы также являются сильным поглотителем углерода растений, тем самым увеличивая градиенты концентрации метаболитов и влияя на корневую экссудацию. Понимание механизмов и влияния экологических стимулов на величину и тип корневой экссудации в конечном итоге улучшит наши знания о процессах, определяющих выбросы CO 2 в почве, функционировании экосистем и способах повышения устойчивости сельскохозяйственного производства.
Поток первичных метаболитов через корневую экссудацию в основном находится на кончике корня, где отсутствие дифференцировки клеток способствует диффузии метаболитов в почву.Мы показываем примеры того, как кончик корня воспринимает изменения концентрации выделяемых метаболитов и преобразует это в сигналы, чтобы изменить рост корня. Растения могут изменять концентрацию метаболитов либо путем управления процессами источника / поглощения, либо путем экспрессии и регулирования переносчиков оттока, тем самым ставя под сомнение идею корневой экссудации как чисто нерегулируемого пассивного процесса. Посредством оттока корневого экссудата растения могут локально увеличивать концентрацию многих общих метаболитов, которые могут служить датчиками и интеграторами состояния питания растений и доступности питательных веществ в окружающей среде.Связанные с растениями микроорганизмы также являются сильным поглотителем углерода растений, тем самым увеличивая градиенты концентрации метаболитов и влияя на корневую экссудацию. Понимание механизмов и влияния экологических стимулов на величину и тип корневой экссудации в конечном итоге улучшит наши знания о процессах, определяющих выбросы CO 2 в почве, функционировании экосистем и способах повышения устойчивости сельскохозяйственного производства.
Введение
Процесс выделения углерода (C) и его адаптивность жизненно важны для растений, чтобы успешно реагировать на изменяющиеся условия окружающей среды.Действительно, максимальное достижение компромисса между инвестициями и доходностью с точки зрения энергии, воды, углерода и питательных веществ в конечном итоге будет определять рост, выживание и взаимодействие растения с его микробиотой. Внешние стрессы, такие как конкуренция, ограничение питательных веществ и / или воды, вызывают серию реакций у растений, которые изменяют распределение углерода, чтобы максимизировать выгоду от ограниченных ресурсов. Множество исследований показали, что подземное распределение углерода в растениях тесно связано с круговоротом воды и питательных веществ (Cheng et al., 2010; McCormack et al., 2015; Гилл и Финци, 2016; Ledo et al., 2018). Тем не менее, главный компонент подземного выделения углерода, а именно процесс корневой экссудации, остается плохо изученным. Остается неясным, почему и как растения вкладывают до 20-40% фотосинтетически фиксированного углерода в корневые экссудаты (Badri and Vivanco, 2009). Современные экологические теории связывают экссудацию корней с пользой для растений через стимуляцию полезных микроорганизмов (например, симбионтов), способствуя усвоению питательных веществ и позволяя распознавать корни и соседние корни (Ortíz-Castro et al., 2009; Dijkstra et al., 2013; Инь и др., 2013; Депюйдт, 2014; Mommer et al., 2016; Meier et al., 2017). Однако хотя некоторые корневые экссудаты, такие как биоактивные вторичные соединения, активно выделяются из корней через энергоемкие первичные или вторичные активные переносчики (Sasse et al., 2018), большинство из них представлены первичными метаболитами (в основном сахарами, аминогруппами). кислоты и органические кислоты), в которых многие исследования предполагают пассивную потерю корня на меристематической вершине корня (McCully and Canny, 1985; Darwent et al., 2003; Jones et al., 2009). В этом контексте возникает несколько фундаментальных вопросов:
1. Каков механизм корневой экссудации первичных метаболитов?
2. Имеют ли растения контроль над этим процессом путем корректировки динамики источников-поглотителей растений и экспрессии переносчиков оттока, и каковы последствия для роста корней?
3. Могут ли растения ощущать концентрацию выделенных и / или переносимых с почвой метаболитов?
4. Участвуют ли эти метаболиты каким-либо образом в добыче питательных веществ через корневую экссудацию?
В этой рукописи мы предлагаем концептуальную основу, основанную на последних достижениях в различных дисциплинах экологии и биологии, связывающих растения с почвенной средой.Здесь мы сосредоточены на первичных метаболитах, которые выделяются в почву (сахара, аминокислоты и органические кислоты) и для которых определенные градиенты концентрации влияют на их корневую экссудацию. Временные концентрации этих метаболитов в кончике корня служат сигналом для восприятия растениями окружающей среды и передачи сигналов между корнями и побегами, чтобы изменить рост корней и выделение углерода. Наша структура предполагает, что корневые экссудаты используются растением, чтобы дополнить функцию переносчиков питательных веществ в определении доступности питательных веществ и в сигнализировании о наличии питательных веществ относительно спроса.Таким образом, этот процесс оптимизирует рост корней, чтобы облегчить эффективный поиск питательных веществ и, возможно, почувствовать comp
Роль функций корней в поглощении кадмия растениями — структурная
- Ресурс исследования
- Исследовать
- Искусство и гуманитарные науки
- Бизнес
- Инженерная технология
- Иностранный язык
- История
- Математика
- Наука
- Социальная наука
Лучшие подкатегории
- Продвинутая математика
- Алгебра
- Основы математики
- Исчисление
- Геометрия
Части вегетативных растений | OSU Extension Service
Внешние структуры растений, такие как листья, стебли, корни, цветы, плоды и семена, известны как органы растения .Каждый орган — это организованная группа тканей, которые работают вместе для выполнения определенной функции. Эти структуры можно разделить на две группы: половых репродуктивных, и вегетативных. Вегетативные частей (рис. 1) включают корни, стебли, почки побегов и листья; они не принимают непосредственного участия в половом размножении. Вегетативные части часто используются при бесполых формах размножения, таких как черенкование, бутонизация или прививка.
Корни
Часто корни упускаются из виду, вероятно потому, что они менее заметны, чем остальная часть растения.Тем не менее, важно понимать корневые системы растений, потому что они оказывают заметное влияние на размер и силу растений, способ размножения, адаптацию к типам почвы и реакцию на культурные обычаи и орошение.
Корни обычно происходят из нижней части растения или черенка. У них есть корневая шляпка, но отсутствуют узлы, и они никогда не имеют листьев или цветов напрямую. Их основные функции — поглощать питательные вещества и влагу, закреплять растение в почве, поддерживать стебель и хранить пищу.У некоторых растений их можно использовать для размножения.
Структура
Внутри корня есть три основные части (рисунок 2):
- Меристема находится на вершине и производит новые клетки; это область деления и роста клеток.
- За меристемой находится зона удлинения . В этой области клетки увеличиваются в размере за счет поглощения пищи и воды. По мере роста они проталкивают корень через почву.
- Зона созревания находится непосредственно под стеблем.Здесь клетки становятся специфическими тканями, такими как эпидермис, кора или сосудистая ткань.
Эпидермис корня — это самый внешний слой клеток (рис. 3). Эти клетки отвечают за поглощение воды и минералов, растворенных в воде. Клетки коры участвуют в перемещении воды из эпидермиса в сосудистой ткани (ксилему и флоэму) и в хранении пищи. Сосудистая ткань расположена в центре корня и проводит пищу и воду.
Внешне важны две области: корневая оболочка и корневые волоски (рис. 2).Корневой колпачок — это крайний край корня. Он состоит из клеток, которые отшелушиваются по мере прорастания корня в почве. Его функция — защита корневой меристемы.
Корневые волоски — это тонкие удлиненные эпидермальные клетки, которые располагаются в небольшой зоне сразу за кончиком корня. Обычно они кажутся прекрасными невооруженным глазом. Их функция заключается в увеличении площади поверхности корня и поглощающей способности. Корневые волоски обычно живут 1-2 дня. При пересадке растения они легко срываются или могут засохнуть на солнце.
Многие корни имеют естественные симбиотические (взаимовыгодные) отношения с определенными грибами, что улучшает способность растения поглощать воду и питательные вещества. Эта полезная ассоциация называется микоризы (гриб + корень).
Виды корней
Есть два основных типа корней: первичный, и боковой. Первичный корень берет начало в нижнем конце зародыша саженца. Если первичный корень продолжает вытягиваться вниз, становится центральным элементом корневой системы и имеет ограниченное вторичное ветвление, он называется стержневым корнем (рис. 4).У гикори и орехов пекан, а также у моркови есть стержневые корни.
Боковой корень или вторичный корень — это боковой корень или корень ветви, который возникает из другого корня. Если первичный корень перестает удлиняться и развиваются многочисленные боковые корни, образуется мочковатая корневая система. Эти боковые корни многократно разветвляются, образуя сеть питающих корней, которая есть у большинства растений.
Некоторые растения, такие как травы, естественным образом дают волокнистую корневую систему . В других случаях отрубание главного корня растения путем его подрезания может стимулировать образование волокнистой корневой системы.Питомники используют эту технику для деревьев, которые естественным образом дают стержневой корень, потому что деревья с компактной мочковатой корневой системой пересаживаются более успешно.
Как растут корни
На раннем этапе развития саженец впитывает питательные вещества и влагу из почвы вокруг прорастающих семян. Полоса удобрений на несколько дюймов в каждую сторону и немного ниже посаженных семян способствует раннему росту большинства пропашных культур.
По мере того, как растение становится устойчивым, количество и распределение его корней сильно влияют на его способность поглощать влагу и питательные вещества.У большинства растений большая часть поглощающих (питающих) корней находится в верхних 12-дюймовых слоях почвы. Почвенная среда в этом регионе обычно лучше всего подходит для роста корней, с хорошим балансом плодородия, влажности и воздушного пространства.
Для роста корней важны следующие факторы:
- Корни в водонасыщенной почве плохо растут и в конечном итоге могут погибнуть из-за недостатка кислорода.
- В рыхлой, хорошо дренированной почве корни проникают намного глубже, чем в тяжелой, плохо дренированной.
- Плотный уплотненный слой почвы может ограничивать или прекращать рост корней.
- Контейнерные растения не только имеют ограниченную зону для роста корней, но также подвержены повреждениям от холода, поскольку ограниченное количество почвы, окружающей их корни, может не обеспечить адекватной изоляции.
- Помимо роста вниз, корни растут сбоку и часто выходят далеко за пределы капельной линии растения. Помните об этой обширной корневой системе, нарушая почву вокруг существующих деревьев и кустарников.
Корни в пищу
Увеличенный корень — съедобная часть некоторых овощных культур. Сладкий картофель — это набухший клубневидный корень; а морковь, пастернак, салсиф и редис — удлиненные стержневые корни.
Стебли
Стебли поддерживают почки и листья и служат проводниками для переноса воды, минералов и пищи (фотосинтатов). Сосудистая система внутри стебля образует непрерывный путь от корня через стебель и, наконец, к листьям. Именно через эту систему движутся вода и пищевые продукты.
Терминология стержня
- Стрельба : Молодой стебель (возрастом менее 1 года) с листьями.
- Веточка : Молодой стебель (возрастом 1 год или меньше), находящийся в зимней стадии покоя (без листьев).
- Ветвь : Стебель возрастом более 1 года, обычно с отходящими от него боковыми стволами.
- Ствол : Главный стебель древесного растения.
Структура
Сосудистая система
Эта система состоит из ксилемы , флоэмы и сосудистого камбия .Его можно рассматривать как водопровод для растений. Xylem трубки проводят воду и растворенные минералы; флоэма пробирок содержат пищу, например сахар. Камбий представляет собой слой меристематической ткани, которая разделяет ксилему и флоэму и постоянно производит новые клетки ксилемы и флоэмы. Эта новая ткань отвечает за увеличение обхвата ствола.
Сосудистый камбий важен для садоводов. Например, ткани на привитом привое и подвое должны выровняться.Кроме того, неосторожная обрезка сорняков может сорвать кору с дерева, тем самым повредить камбий и вызвать гибель дерева.
Сосудистые системы однодольных и двудольных различаются (рис. 5). Хотя оба содержат ксилему и флоэму, эти структуры устроены в каждой по-разному. У однодольных ксилема и флоэма объединены в пучки, которые рассредоточены по стеблю. У двудольных растений сосудистая система называется непрерывной, потому что она образует кольца внутри стебля. Кольцо флоэмы находится рядом с корой и со временем становится частью коры зрелых древесных стеблей.Ксилема образует внутреннее кольцо. У древесных растений это называется заболонью и сердцевиной.
Различие сосудистых систем однодольных и двудольных представляет практический интерес для садоводов, поскольку некоторые гербициды действуют только на одну группу. Например, 2,4-Д убивает только растения с непрерывной сосудистой системой (двудольные). С другой стороны, неселективные гербициды (например, глифосат) убивают растения независимо от типа их сосудистой системы.
Узлы
Узел — это участок на стебле, где расположены почки (Рисунок 6).Это место высокой клеточной активности и роста, где маленькие бутоны развиваются в листья, стебли или цветы. При обрезке важно найти узлы растения. Как правило, вы хотите сделать обрезку чуть выше, но не слишком близко к узлу. Такая обрезка побуждает бутоны на этом узле начать развитие и в конечном итоге сформировать новые стебли или листья.
Область между двумя узлами называется межузлом . Его длина зависит от многих факторов, в том числе от генетики.Несколько других факторов также могут влиять на длину междоузлия:
- Пониженное плодородие почвы уменьшает длину междоузлий, в то время как внесение удобрений с высоким содержанием азота может значительно увеличить ее.
- Недостаток света увеличивает длину междоузлий и делает стержень веретенообразным. Эта ситуация известна как растяжение, или этиоляция , и часто возникает у всходов, выращенных в помещении, и у комнатных растений, которые не получают достаточно солнечного света.
- Длина междоузлия также меняется в зависимости от сезона.Рост в начале сезона имеет длинные междоузлия, в то время как рост в конце сезона обычно имеет гораздо более короткие междоузлия.
- Если энергия стебля разделяется между тремя или четырьмя боковыми стеблями или направляется на рост и развитие плодов, длина междоузлий сокращается.
- Вещества, регулирующие рост растений, и гербициды также могут влиять на длину междоузлий.
Типы стеблей
Стебли могут быть длинными, с большими расстояниями между листьями и почками (например, ветки деревьев, побеги клубники) или сжатыми, с небольшими расстояниями между почками или листьями (например, ветви деревьев, побеги клубники).г., кроны клубники, шпоры плодовых, африканские фиалки). Хотя стебли обычно растут над землей, иногда они растут под землей в виде корневищ, клубней, клубнелуковиц или луковиц. Все стебли должны иметь бутоны или листья, чтобы их можно было классифицировать как ткань стебля.
Стволы специализированные надземные
У некоторых растений есть специализированные надземные стебли, известные как кроны , шпоры или столоны (рис. 7). Короны (клубника, одуванчики и африканские фиалки) представляют собой сжатые стебли с листьями и цветами на коротких междоузлиях. Шпоры — короткие, короткие боковые штанги, которые отходят от основного штока. Это плодовые стебли груши, яблони и вишни. Если провести сильную обрезку рядом с плодоносящими отростками, они могут вернуться к неплодоносящим стеблям, тем самым исключив потенциальный урожай плодов в этом году.
Столоны — это мясистые или полудревесные, удлиненные, горизонтальные стебли, которые часто лежат вдоль поверхности почвы. Побеги клубники — это столоны с небольшими листьями в узлах. Из этих узлов развиваются корни, и образуется дочернее растение.Этот тип вегетативного размножения — простой способ увеличить размер клубники. Растения-пауки также производят столоны, которые в конечном итоге могут стать совершенно новыми растениями.
Специализированные подземные стволы
Клубни картофеля, корневища ириса и луковицы тюльпанов — это подземные стебли, на которых хранится корм для растения (рис. 8). Иногда бывает трудно отличить корни от стеблей, но верный способ — поиск узлов. Стебли имеют узлов ; корней нет.
У картофеля клубни , например, «глазки» на самом деле являются узлами стебля, и каждый глазок содержит скопление почек. При выращивании картофеля из кусков семян важно, чтобы каждый кусочек имел хотя бы один глазок и был размером с мяч для гольфа, чтобы у него было достаточно энергии для раннего роста побегов и корней.
Корневища напоминают столоны, потому что они растут горизонтально от растения к растению. Некоторые корневища сжатые и мясистые (например, радужная оболочка), в то время как другие тонкие и имеют удлиненные междоузлия (например, радужная оболочка).г., полевник). Johnsongrass — коварный сорняк, главным образом из-за способности его корневищ к распространению.
Тюльпаны, лилии, нарциссы и лук дают луковиц , которые представляют собой укороченные сжатые подземные стебли, окруженные мясистыми чешуйками (листьями), которые охватывают центральный бутон на кончике стебля. В ноябре можно разрезать луковицу тюльпана или нарцисса пополам и увидеть все части цветка в миниатюре.
После цветения луковичного растения его флоэма переносит запасы пищи с листьев на чешуйки луковицы.Когда луковица начинает расти весной, она использует накопленную пищу. По этой причине важно не удалять листья нарциссов, тюльпанов и других растений, производящих луковицы, до тех пор, пока они не пожелтеют и не засохнут. К тому времени они закончили производство корма, который будет использоваться для цветения в следующем году.
Есть два типа ламп: оболочковые и несвязанные (Рисунок 8). Tunicate Луковицы (например, нарциссы, тюльпаны и лук) имеют тонкую бумажную оболочку, которая на самом деле представляет собой модифицированный лист.Это помогает защитить луковицу от повреждений во время перекопки и от высыхания после выхода из почвы. Нераздельные луковицы (например, лилии) не имеют бумажного покрытия. Они очень подвержены повреждениям и высыханию, поэтому обращайтесь с ними очень осторожно.
Клубнелуковицы — еще один вид подземных стеблей. Хотя луковицы и клубнелуковицы состоят из стволовой ткани, они не совпадают. Клубнелуковицы имеют форму луковиц, но не содержат мясистых чешуек. Клубнелуковица — это твердый вздутый стебель с сухими чешуевидными листьями.Гладиолусы и крокусы производят клубнелуковицы.
Некоторые растения (например, клубневые бегонии и цикламены) производят модифицированный подземный стебель, называемый клубневым стеблем . Эти стебли короткие, плоские и увеличенные. Почки и побеги возникают сверху (кроны), а снизу растут мочковатые корни.
Другие растения (например, георгины и сладкий картофель) производят подземные запасающие органы, называемые клубневые корни . которые часто путают с луковицами и клубнями. Однако это ткань корня, а не ткань ствола, и у нее нет ни узлов, ни междоузлий.
Стебли и размножение
Стебли часто используются для вегетативного размножения растений. Использование участков надземных стеблей, содержащих узлы и междоузлия, является эффективным способом размножения многих декоративных растений. Эти стеблевые черенки дают корни и, в конечном итоге, новые растения.
Подземные стебли также являются хорошими тканями для размножения. Можно корневища разделить на части; удалите с их родителей мелкие луковицы или кормели; и разрезать клубни на части, содержащие глазки и узлы.Все эти ткани дадут новые растения.
Виды растений и их стебли
Деревья обычно имеют один, но иногда и несколько основных стволов, которые в зрелом возрасте обычно достигают высоты более 12 футов. Напротив, кусты обычно имеют несколько основных стеблей, которые в зрелом возрасте обычно достигают менее 12 футов в высоту.
Большинство фруктовых деревьев, декоративных деревьев и кустарников имеют древесные стебли. Эти стебли содержат относительно большое количество затвердевшей ткани ксилемы в центральном ядре (сердцевине или заболони).
Травянистые или сочные стебли содержат лишь немного ткани ксилемы и обычно живут только один вегетационный период. У многолетних растений новые стебли травянистых растений развиваются из кроны (поверхности раздела корень-стебель) каждый год.
Трость (рис. 9а) — это стержень с относительно большой сердцевиной (центральная ткань, придающая силу). Обычно они живут всего 1-2 года. Примеры растений с тростником: розы, виноград, ежевика и малина. Для производства фруктов важно знать, какие стебли обрезать, как обрезать и когда обрезать.
Виноградная лоза (рис. 9b) — это растение с длинными висящими стеблями. Некоторые виноградные лозы растут по земле, в то время как другие должны поддерживаться другим растением или строением. Вьющиеся лозы окружают конструкцию для поддержки. Некоторые кружатся по часовой стрелке (например, хмель и жимолость), а другие — против часовой стрелки (например, бобы и лоза голландца). Вьющиеся лозы поддерживаются либо воздушными корнями (например, английский плющ и ядовитый плющ), либо тонкими усиками, которые окружают поддерживающий объект (например, английский плющ и ядовитый плющ).g., огурцы, тыквы, виноград и пассифлоры) или усиками с липкими кончиками (например, вирджиния и японская лиана).
Стебли в пищу
Съедобная часть некоторых культурных растений, таких как спаржа и кольраби, представляет собой увеличенный сочный стебель. Съедобные части брокколи состоят из стеблевой ткани, цветочных почек и нескольких маленьких листьев. Съедобный клубень картофеля — это мясистый подземный стебель. И хотя название говорит об обратном, съедобная часть цветной капусты на самом деле представляет собой разросшуюся стеблевую ткань.
Почки
Бутон — это неразвитый побег, из которого растут листья или части цветка. На почках деревьев и кустарников умеренного пояса обычно образуется защитный внешний слой из мелких кожистых чешуек. Однолетние растения и травянистые многолетники имеют голые бутоны с зелеными, немного сочными внешними листьями.
Почки многих растений требуют выдержки в течение определенного количества дней при температуре ниже критической, прежде чем возобновить рост весной. Этот период, часто называемый покоем, у разных растений различен.Например, форзиция требует относительно короткого периода отдыха и растет при первых признаках теплой погоды. С другой стороны, многим сортам персиков требуется от 700 до 1000 часов при температуре ниже 45 ° F. Во время покоя спящие бутоны могут выдерживать очень низкие температуры, но после того, как период покоя насыщен, они более подвержены повреждению холодными температурами или морозами.
Листовая почка состоит из короткого стебля с зародышевыми листьями. Листовые почки часто менее пухлые и более заостренные, чем цветочные почки (рис. 10а).Бутон цветка состоит из короткого стебля с зародышевыми частями цветка. В случае плодовых культур цветочные почки иногда называют плодовыми. Однако эта терминология неточна; Хотя цветы могут развиваться в плоды, это может не произойти из-за неблагоприятных погодных условий, отсутствия опыления или других неблагоприятных обстоятельств.
Расположение
Почки названы в честь их расположения на стебле (рис. 10b). Терминал почки расположены на вершине (кончике) стебля.Боковые ( пазушные, ) почки расположены по бокам стебля и обычно возникают там, где лист встречается со стеблем (пазуха , ). В некоторых случаях пазуха содержит более одной почки.
Придаточные зародыши () возникают в других местах, кроме терминального или подмышечного положения. Они могут развиваться из корней, стебля , междоузлия , края листовой пластинки или каллусной ткани на отрезанном конце стебля или корня. Придаточные почки позволяют черенкам стебля, листьев и корней развиваться в совершенно новые растения.
Почки в пищу
Увеличенные почки или части почек составляют съедобную часть некоторых садовых культур. Кочанная капуста и кочанный салат — примеры необычайно крупных верхушечных почек. Сочные пазушные почки — съедобная часть брюссельской капусты. В случае шарообразного артишока съедается мясистая базальная часть прицветников бутона вместе с его твердым стеблем. Брокколи — важнейшее садовое растение со съедобными цветочными бутонами. В этом случае поедаются части стебля, а также небольшие листья, связанные с цветочными бутонами.
Листья
Функции и устройство
Основная функция листьев — поглощение солнечного света для производства растительных сахаров посредством процесса, называемого фотосинтез . Поверхности листьев сплющены, чтобы обеспечить большую площадь для эффективного поглощения света. Лезвие представляет собой расширенную тонкую структуру по обе стороны от средней жилки и обычно является самой большой и заметной частью листа (рис. 11).
Лист удерживается на расстоянии от стебля стеблевидным отростком, называемым черешком , а основание черешка прикреплено к стеблю в узле .Черешки различаются по длине или могут отсутствовать полностью, в этом случае листовая пластинка описывается как сидячая, или бесстебельная.
Узел, где черешок встречается со стеблем, называется пазухой листа . В пазухе содержатся отдельные почки или скопления почек, называемые пазушными почками. Они могут быть активными или бездействующими; в правильных условиях они разовьются в стебли или листья.
Листовая пластинка состоит из нескольких слоев (Рисунок 12a и Рисунок 12b). Сверху и снизу находится слой толстых твердых клеток, называемый эпидермисом .Его основная функция — защищать другие слои ткани листа. Расположение клеток эпидермиса определяет текстуру поверхности листа. У некоторых листьев, например у африканской фиалки, есть волоски (опушение , ), которые являются продолжением эпидермальных клеток, благодаря которым листья кажутся бархатными.
Кутикула является частью эпидермиса. Он образует восковой слой, называемый кутин , который защищает лист от обезвоживания и болезней. Количество кутина на листе увеличивается с увеличением интенсивности света.По этой причине, перемещая растения из тени на солнечный свет, делайте это постепенно в течение нескольких недель. Постепенное воздействие солнечного света позволяет нарастить слой кутина и защитить листья от быстрой потери воды или солнечного ожога.
Восковый кутин также отталкивает воду. По этой причине многие пестициды содержат аэрозольные добавки, которые помогают продукту прилипать или проникать через слой кутина.
Специальные эпидермальные клетки, называемые замыкающими клетками, открываются и закрываются в ответ на внешние раздражители, такие как изменения погоды и света.Они регулируют прохождение воды, кислорода и углекислого газа в лист и из него через крошечные отверстия, называемые устьицами . У большинства видов большинство устьиц расположено на нижней стороне листьев.
Условия, при которых растения теряют много воды (высокая температура, низкая влажность), стимулируют закрытие замыкающих клеток. В мягкую погоду они остаются открытыми. Сторожевые клетки также закрываются при отсутствии света.
Между верхним и нижним эпидермисом находится мезофилл .Он разделен на плотный верхний слой (палисадный мезофилл) и нижний слой, содержащий много воздушного пространства (губчатый мезофилл). Внутри клеток мезофилла находятся хлоропластов , где происходит фотосинтез.
Виды листьев
Есть много видов листьев растений. Наиболее распространенные и заметные листья называются листвой и являются основным местом фотосинтеза. Однако существует множество других видов модифицированных листьев:
- Чешуйчатые листья (катафиллы) находятся на корневищах и почках, которые они закрывают и защищают.
- Семядоли ( семядолей ) встречаются на зародышевых растениях. В них хранится корм для развивающегося саженца.
- Колючки и усики , такие как те, что встречаются у растений барбариса и гороха, защищают растение или помогают поддерживать его стебли.
- Листья для хранения , например, на луковичных и суккулентных растениях, служат для хранения продуктов.
- Прицветники часто ярко окрашены. Например, эффектные структуры на кизилах и пуансеттиях представляют собой прицветники, а не лепестки.
Расположение
Сосудистые пучки ксилемы и флоэмы проходят от стебля через черешок до листовой пластинки в виде жилок. Термин жилкование относится к тому, как жилки распределены в лезвии. Существует два основных типа жилкования: с параллельными жилками и с сетчатыми жилками (рисунок 13).
В листах с параллельными прожилками многочисленные прожилки идут по существу параллельно друг другу и соединяются сбоку мельчайшими прямыми прожилками.Листья с параллельными жилками чаще всего встречаются на однодольных растениях. Наиболее распространенный тип параллельных жилок встречается у растений семейства злаковых, жилки которых проходят от основания листа к его вершине. Другой тип параллельного жилкования встречается у таких растений, как банан, калла и пикерник, жилки которых идут сбоку от средней жилки.
В листьях с сетчатыми прожилками (также называемых с сетчатыми прожилками ) жилки ответвляются от главного ребра или ребер и разделяются на более тонкие прожилки.Затем эти прожилки объединяются в сложную сеть. Эта система переплетенных жилок делает лист более устойчивым к разрыву, чем структура с параллельными жилками. Листья с сетчатыми прожилками встречаются у двудольных растений.
Сетчатое жилкование может быть перистым или пальчатым. У перистого жилкования жилки проходят латерально от средней жилки до края (например, у яблок, вишни и персиков). В жилковании пальчато- основные жилки выходят наружу, как ребра веера, от основания листовой пластинки (e.г., виноград и клены).
Листья как идентификаторы растений
Листья используются для идентификации растений. Жилкование листа (Рисунок 13), форма лезвия и края (Рисунки 14 и 15), а также форма вершины и основания (Рисунок 16) могут быть важными отличительными характеристиками.
Тип листа (Рисунок 17) также важен для идентификации. Есть два типа листов: простые и составные . В простых листах листовая пластина представляет собой единый сплошной блок. Сложные листья состоят из нескольких отдельных листочков, растущих на одном черешке. Некоторые листья двояко сложны. Тип листа может сбивать с толку, потому что простой лист с глубокими лопастями может выглядеть как сложный.
Расположение листьев вдоль стебля также используется при идентификации растений (рис. 18). Есть четыре типа расположения створок:
- Противоположные листья расположены поперек стебля друг от друга, по два листа на каждом узле.
- Альтернативные (спиральные) листья расположены попеременно вдоль стебля, только по одному листу в каждом узле.
- Мутовчатые листья расположены по кругу вдоль стебля.
- Рошеобразные листья расположены в виде розетки вокруг стебля с очень короткими узлами.
Листья в пищу
Листовая пластинка является основной съедобной частью некоторых садовых культур, в том числе чеснока, капусты, одуванчиков, эндивий, капусты, листового салата, горчицы, петрушки, шпината, мангольда и другой зелени. Съедобная часть лука-порея, лука и флорентийского фенхеля представляет собой группу оснований мясистых листьев.Черешок — это съедобный продукт сельдерея и ревеня.
Регулирующий кальций для растений для лучшего питания и устойчивости к стрессу
Внешний Ca 2+ Добавка помогает растениям восстанавливаться после стресса. В этой статье рассматриваются генетические методы увеличения Ca 2+ для повышения устойчивости растений к стрессу и повышения их питательной ценности. Транспорт Ca 2+ должен тщательно контролироваться, чтобы минимизировать колебания в цитозоле, обеспечивая при этом как структурную поддержку новых клеточных стенок и мембран, так и внутриклеточные запасы Ca 2+ для передачи сигналов.Неясно, как это достигается в меристемах, удаленных от активной транспирации — движущей силы движения Ca 2+ в побеги. Меристемы имеют высокий уровень кальретикулина (CRT), который связывает 50-кратный избыток Ca 2+ и может способствовать транспорту Ca 2+ между клетками через плазмодесматальный ER. Трансгены, основанные на высокомощном Ca 2+ -связывающем C-домене CRT1 , увеличили общий Ca 2+ растений на 15-25%, а также повысили устойчивость к абиотическому стрессу.Эти результаты сравнивают со сверхэкспрессией sCAX1 , которая не только увеличивала общий Ca 2+ до 3 раз, но также вызывала симптомы дефицита Ca 2+ . Совместная экспрессия sCAX1 и CRT1 разрешила симптомы и привела к высоким уровням Ca 2+ без добавления Ca 2+ . Эти результаты предполагают важную роль ER Ca 2+ в толерантности к стрессу и передаче сигналов и демонстрируют возможность использования белков, модулирующих Ca 2+ , для улучшения как агрономических, так и пищевых свойств.
1. Введение
Растения чувствуют раздражители окружающей среды и реагируют на них, используя сети сенсоров, вторичных мессенджеров, киназ и факторов транскрипции для регулирования экспрессии генов и адаптации к новым условиям. Ca 2+ , пожалуй, самый известный вторичный посредник, но он также необходим для правильной структуры клеточной стенки и целостности мембраны [1]. Хотя Ca 2+ присутствует в относительно высоких концентрациях (0,1–80 мМ) в клеточных стенках и органеллах, цитоплазматические уровни Ca 2+ поддерживаются на уровне ~ 100 нМ [2–4].Для передачи сигнала в растениях требуется способность мобилизовать и изолировать Ca 2+ как из внутренних, так и из внешних запасов Ca 2+ . Поскольку как дефицит, так и высокие концентрации Ca 2+ вызывают локальную гибель клеток, транспорт Ca 2+ по всему растению должен строго регулироваться [5, 6].
Растения, выращенные в условиях дефицита Ca 2+ , более восприимчивы к патогенам растений и демонстрируют снижение роста апикальных меристем, хлоротичных листьев и разрушение клеточной стенки, что приводит к размягчению тканей [2].Но добавление Ca 2+ не только облегчает эти симптомы, но и способствует росту растений за счет увеличения длины корней и помогает им выдерживать стресс или восстанавливаться после него [7–14]. Добавка Ca 2+ также используется для улучшения характеристик плодов и может замедлять вызванное этиленом старение [15]. Эта информация не нова, в отчете Science , опубликованном более 40 лет назад, описывается эффект 1 мМ Ca 2+ в предотвращении тяжелой токсичности NaCl в бобах [16].
Точные эффекты внеклеточного Ca 2+ на систему растений, вероятно, будут сложными, потому что Ca 2+ выполняет несколько ролей и потому что разные растения демонстрируют разные ответы на дополнительный Ca 2+ . Например, у большинства растений внеклеточный Ca 2+ снижает накопление Na + , что снижает солевой стресс. Но у некоторых растений (например, кукурузы) уровни Na + остаются постоянными, но благотворное влияние на рост растений все еще очевидно [17].Добавка Ca 2+ в некоторые растения, такие как рис, не оказывает видимого влияния на солеустойчивость [18] (см. [19]). Добавление K + либо не оказывает никакого эффекта, либо вредно [20].
В недавнем отчете было обнаружено, что раствор Ca 2+ полезен при распылении непосредственно на листья чайных растений, подвергшихся стрессу от засухи [21]. Какую пользу растениям приносит простое распыление Ca 2+ на листья? Почему заводы не придумали, как увеличить свои собственные запасы, поскольку Ca 2+ легко доступен в большинстве сред? Кроме того, является ли это частью того, что делает некоторые растения «сорными»? Есть ли препятствия для эффективного переноса на большие расстояния Ca 2+ ? Можем ли мы разработать «обходной» или альтернативный механизм для транспорта Ca 2+ , чтобы помочь им оправиться от стресса или даже предотвратить повреждение в первую очередь?
Чтобы понять, как внеклеточный Ca 2+ приносит пользу растениям, необходимо больше узнать о функции и подвижности Ca 2+ как на клеточном, так и на уровне всего растения.Как только мы поймем различные роли Ca 2+ , как Ca 2+ секвестрируется и транспортируется в растении, высвобождается для передачи клеточных сигналов, а затем быстро изолируется от пагубных взаимодействий, тогда мы можем начать думать о пересмотре или адаптируя некоторые из его путей. В этой статье будет представлен краткий обзор регулируемых путей и функций Ca 2+ на всем растении с целью выявления потенциальных стратегий для создания дополнительных ионов Ca 2+ в растворимых запасах растений, чтобы они были легко доступны для сигнализация и рост.Есть надежда, что этот подход может быть частью стратегии по созданию более питательных сельскохозяйственных культур, которые также будут более устойчивы к стрессу.
2. Ca 2+ Хранение и передача сигналов
Принято считать, что Ca 2+ , один из самых распространенных минералов на Земле, превратился в сигнальную молекулу из-за двойной потребности клетки. для растворимого фосфата и Ca 2+ и их склонность к выпадению в осадок в виде нерастворимой соли [22].Фосфат также играет важную роль в передаче сигнала, но его роль в качестве промежуточного звена энергии требует присутствия в цитоплазме [23]. У растений метаболические пути, в которых используется АТФ, обнаруживаются в основном в цитоплазме и отделены от запасов Ca 2+ , которые находятся в основном в апопласте, вакуоли и эндоплазматическом ретикулуме (ER) и в меньшей степени в митохондриях. хлоропласты и ядро [4]. В клетках животных и на ранних этапах линии растений ER был основным источником Ca 2+ , и его высвобождение контролировалось другим вторичным мессенджером, инозитол (1,4,5) трифосфатом (IP 3 ), через активация ER-локализованных рецепторов IP 3 [24, 25].Подобные рецепторы IP 3 не были обнаружены у растений; однако фосфоинозитидный путь у растений консервативен [26–29]. Члены фосфоинозитидного сигнального пути демонстрируют регуляцию транскрипции с помощью стимулов окружающей среды и развития у Arabidopsis [30], а высвобождение Ca 2+ с помощью IP 3 является консервативным [31, 32].
Помимо апопласта, ER и вакуоли являются основными и метаболически значимыми источниками клеточного Ca 2+ [33–35].Цитозольные уровни Ca 2+ колеблются и контролируются системой локализованных на мембране насосов Ca 2+ и каналов Ca 2+ , расположенных в плазмалемме, вакуоли и ER [4, 5, 36]. Электрохимический потенциал Ca 2+ для проникновения в цитоплазму через плазматическую мембрану, рассчитанный Сполдингом и Харпером, составляет около -52 кДж / моль [22]. Следовательно, Ca 2+ может проникать в клетки пассивно через ионные каналы, но требует энергии для откачки из цитоплазмы.Хотя энергетически невыгодно, удаление Ca 2+ происходит быстро и эффективно, что приводит к 1000-кратным и большим различиям [Ca 2+ ] между цитозолем и окружающими органеллами и апопластом [37].
В отличие от животных систем, мутации в транспортных белках Ca 2+ часто не вызывают драматических фенотипов [22], предполагая, что растения более устойчивы к цитозольному Ca 2+ или что у них есть перекрывающиеся и дублирующие системы. Это затрудняет сопоставление электрофизиологических экспериментов с генетикой, чтобы точно определить, какие каналы Ca 2+ функционируют в передаче сигналов (или хранении) и когда.Было показано, что Ca 2+ высвобождается из ER и, возможно, других мембран с помощью cADP-рибозы, метаболита NAD + , аналогично тому, что происходит в клетках животных, более десяти лет назад [34], однако теперь кажется Ясно, что циклические нуклеотид-управляемые каналы (CNGC) обнаруживаются в плазмалемме [48]. Один из немногих белков, которые действительно имеют фенотип, фенотип cngc2 , подобен cax1 / cax3 (см. Раздел 3), что позволяет предположить, что он играет важную роль в обеспечении несигнального входа Ca 2+ в клетки листа. [49].В Arabidopsis имеется 20 генов CNGC и еще 20 генов, которые кодируют глутаматные рецепторные каналы (GLR), другой тип канала Ca 2+ , обнаруженный в плазмалемме [48]. Третий тип канала, двухпоровый канал Ca 2+ (TPC1), был сначала идентифицирован как белок плазмалеммы, но теперь известно, что он локализован на мембране тонопласта.
Есть две основные группы белков, которые участвуют в удалении Ca 2+ из цитоплазмы [50]. Автоингибирующая АТФаза Ca 2+ (ACA) использует энергию АТФ для перекачки Ca 2+ из цитоплазмы в органеллы, такие как вакуоль и ER.Вторая группа белков функционирует как антипортеры и называется белками CAtion eXchange (CAX) и находится на мембране тонопласта. CAX обменивает два протона на один Ca 2+ , используя энергию градиента протонов для подавления цитоплазматических сигналов Ca 2+ [51].
2.1. Calcium Signatures
Цитоплазматическое увеличение Ca 2+ в ответ на высокие концентрации соли было отмечено, по крайней мере, 25 лет назад у растений [52], но специфичность передачи сигналов Ca 2+ все еще недостаточно изучена.Существуют две неисключительные модели того, как Ca 2+ функционирует в качестве второго мессенджера. Модель сигнатуры Ca 2+ утверждает, что информация закодирована в форме, продолжительности и частоте переходных процессов Ca 2+ и разнообразии клеточных хранилищ Ca 2+ , все из которых может способствовать формированию микродоменов, которые поддерживают и реагируют на локализованные изменения Ca 2+ [4, 53]. Эти локализованные изменения специфичны для индуцирующего стимула и приводят к специфическим изменениям белков, модулируемых Ca 2+ , и их мишеней [5, 39, 54–56].Вторая модель предполагает, что переходные процессы Ca 2+ функционируют как простой бинарный переключатель, включается или выключается, и именно датчик Ca 2+ (белок, модулированный Ca 2+ ) связывает различные стимулы с адаптивный ответ [22].
Наиболее изученные примеры передачи сигналов, опосредованной Ca 2+ , включают открытие замыкающих клеток, клубеньков и рост кончиков поляризованных структур, таких как пыльцевые трубки [57–66]. Сообщалось также о специфических сигнатурах Ca 2+ , например, в ответ на различные химические вещества в корне (алюминий, глутаминовая кислота и АТФ [67]) и, на уровне всего растения, в ответ на озон [68] .Примеры других стимулов, которые вызывают временное повышение концентраций Ca 2+ в цитозоле, включают прикосновение, холодовой шок, тепловой шок, оксидативный стресс, аноксию, гипоосмотический шок, соленость, ранение, гравитацию и инфекцию патогенами [37, 56, 69 –81]. Сигналы развития, включая оплодотворение, старение, оплодотворение и созревание, также включают белки, регулируемые Ca 2+ [82–88].
Имеются данные о тканеспецифических различиях в потоке Ca 2+ в ответ на один и тот же стимул, например, солевой стресс.Солеустойчивость — сложный признак, включающий ответы на клеточные осмотические и ионные стрессы и связанные с ними вторичные стрессы (например, окислительный стресс) [89, 90]. В корнях наблюдается двухфазное временное увеличение цитозольного Ca 2+ после воздействия острого солевого стресса [73]. В отличие от холодового шока, который ограничен областями около меристемы корня, солевой шок увеличивает цитозольный Ca 2+ по всему корню [91]. Чтобы различить тканеспецифические различия в потоке Ca 2+ , различные трансгенные растения, трансформированные геном, кодирующим экворин (репортерный ген для Ca 2+ ), нацелены на цитоплазму эпидермиса, энтодермы или перицикла корней арабидопсиса.