
Щавель — многолетнее, двудомное травянистое растение семейства гречишные (Polygonaceae). В Российской Федерации и Украине выращивают в качестве овощного растения щавель обыкновенный, или кислый (Rumex acetosa L.) и частично щавель шпинатный (Rumex patientia L.). Щавель кислый живет до 8—10 лет, на продовольственные цели его выращивают 1,5—2 года, на семенные — 4—5 лет. Куст растения компактный. От головки корня отрастает розетка листьев на укороченных побегах, что позволяет размножать щавель вегетативно. Розетка раскидистая или компактная, приподнятая или прямостоячая. Цветоносные стебли появляются на второй год в центре каждой розетки. Новые боковые почки развиваются по периферии, с возрастом середина куста становится пустой, а розетки располагаются по кругу. Стебель прямостоячий, высотой 70—100 см, ребристый, облиственный, ветвящийся только в верхней части соцветия. В нижней части его листья черешковые, в верхней до соцветия — сидячие. Растение раздельнополое. Генеративные стебли с женскими цветками обычно выше, чем с мужскими; количество их в 4—5 раз больше, чем мужских. Окраска цветоносного стебля зеленая, при созревании семян — бурая. Листья довольно мясистые, прикорневые и нижние стеблевые — длинночеренковые, яйцевидно-продолговатые со стреловидным или копьевидным основанием, на верхушке тупо заостренные или заостренные, голые. Длина пластинки листа 15—22 см, ширина — 5—12 см, диаметр черешка — 3—4 мм. Окраска листа зеленая с темно-, светло-или желтовато-зеленым оттенком. Внутренние листочки околоцветника розовые, красные, желтоватые, смешанной окраски. У мужских цветков листочки околоцветника продолговато-овальные, внутренние немного крупнее наружных, все направлены кверху, опадающие. У женских цветков наружные листочки околоцветника при плодах перепончатые, отгибаются, прижаты к цветоножке, у основания немного вздуты, иногда сросшиеся между собой. Головка корня имеет большое количество почек, из которых вырастают листья. По мере уборки урожая пробуждаются спящие почки, из которых формируются новые листья. Соцветие метельчатое. Сначала оно зеленое, затем приобретает красноватую, а при созревании семян — бурую окраску. Соцветие рыхлое, узкое, ветвление слабое, цветки собраны в негустые кисти. Цветки мелкие, однополые, изредка обоеполые; окраска венчика красновато-желтоватая, иногда розоватая. Цветение у щавеля в зависимости от климатических условий длится с мая до августа. Опыление цветков происходит при помощи ветра и насекомых. Семена созревают через 65—-75 суток после появления цветоносных стеблей. С одного участка щавеля семена можно убирать 3—4 года, но лучший урожай и посевные качества их получают на 2— 3-й год жизни растений. Плод — сухой, односемянный, остроконечный, нераскрывающийся, трехгранный, мелкий орешек (семянка). Поверхность блестящая, гладкая. Окраска темно-коричневая, по граням — беловатая. Длина семени 1,5—2,7 мм, ширина — 1,0—1,6 мм, толщина — 1,0—1,4 мм. Масса 1000 семян — 0,8—1,2 г. Семена сохраняют всхожесть 3—4 года. При семеноводстве пространственная изоляция между видами и сортами должна быть не менее 2 км, при наличии лесополос — 1 км. Корень стержневой, разветвленный, тонкий, в верхнем слое почвы образует корневище. Верхняя часть корня диаметром 10—15 мм, на глубине 25 см — 2—3 мм. Боковые ветви корня тоже тонкие. Новые побеги на головке корня с розетками листьев имеют свою мочковатую корневую систему из тонких боковых корней длиной 10—15 см. Основная масса корней расположена в пахотном слое (0— 25 см), отдельные корни уходят на глубину до 35—40 см. Корни щавеля накапливают большое количество питательных веществ. Благодаря этому они успешно переносят зимние холода. Однако при отсутствии снежного покрова при —15…—20°С наблюдается гибель растений. Известно более 150 видов щавеля. Большинство их произрастает в умеренных широтах северного полушария. На территории СНГ можно встретить около 40 видов. Все они преимущественно дикорастущие. Среди культивируемых форм большое внимание заслуживает щавель шпинатный, или английский (Rumex patientia L.). На территории СНГ можно встретить около 40 видов. Все они преимущественно дикорастущие. Среди культивируемых форм большое внимание заслуживает щавель шпинатный, или английский (Rumex patientia L.).Щавель шпинатный больше распространен на Кавказе, в странах Центральной Европы и Средиземноморья, в Малой Азии, Иране, Монголии. В Европе известен как шпинат многолетний, шпинат английский, шпинат озимый. От щавеля кислого отличается более крупными эллипсовидными листьями на длинных черешках, шириной до 9 см, по краю волнистыми, меньшим содержанием щавелевой кислоты, более высоким — яблочной, лимонной и аскорбиновой, слабокислым вкусом и скороспелостью. Щавель размножают семенами, при селекционной работе применяют и вегетативный способ. При высеве семян ранней весной он дает хороший урожай зелени уже в первый год. К уборке урожая приступают, когда растения развивают 4—5 настоящих листьев или через 45—50 суток после всходов. Щавель холодостойкое и морозостойкое растение. Хорошо зимует. Семена начинают прорастать при температуре +1…+2°С. Однако первые всходы появляются при температуре +18…+20°С. Оптимальной температурой для роста и развития растений является + 16…+18°С. Листья могут отрастать до самых морозов; при температуре выше +25°С рост их замедляется, они грубеют, урожай снижается. Нераспустившиеся почки выдерживают весенние заморозки до —7…—10°С. Техническая спелость зелени наступает через 14—20 суток после оттаивания почвы. Щавель хорошо сохраняется зимой под снежным покровом. Чувствительны к низким температурам растения поздних сроков сева, которые накапливают в корневищах мало питательных веществ. В первый год жизни растения щавеля предъявляют высокие требования к освещенности. Особенно чувствительны к недостатку освещения всходы. В дальнейшем, когда в корнях больше накапливается питательных веществ, временный недостаток света не оказывает существенного влияния на продуктивность растений. Поэтому взрослые растения более теневыносливы. Щавель — влаготребовательное растение, но плохо растет на низких, заболоченных участках с уровнем грунтовых вод выше 1 м от поверхности почвы. В засушливые весны рост приостанавливается, листья грубеют и отрастают более мелкими. При недостатке влаги, кроме того, ускоряются процессы развития. В результате начинается преждевременное и обильное стеблевание растений. На рост стеблей расходуется большое количество питательных веществ, формирование листьев задерживается и снижается урожай. Для нормального роста и развития требуется высокая влажность почвы и воздуха. Во всех почвен-но-климатических зонах щавель размещают на орошаемых землях. Оптимальную влажность почвы необходимо поддерживать поливами на уровне 80% НВ. Поливы проводят после каждой срезки листьев, что ускоряет пробуждение спящих почек и способствует быстрому отрастанию новых листьев. К почвенному питанию щавель предъявляет повышенные требования. Под его посевы отводят высокоплодородные, легко прогреваемые, легко- и среднесуглинистые, хорошо окультуренные почвы, содержащие не менее 4% гумуса. Подпочвенный слой должен быть проницаемым для воды. На участках с водонепроницаемой подпочвой, где застаиваются весенние воды, щавель медленно трогается в рост и дает листья с горьковатым привкусом, а также в этих условиях он менее устойчив к весенним заморозкам. Тяжелые глинистые почвы образуют плотную корку, из-за чего всходы получаются невыравненными или изреженными и развитие их замедляется. Для щавеля одинаково подходят как кислые почвы (рН 4,5—5,5), так и нейтральные. Листья щавеля отрастают ранней весной, когда в почве недостаточно азота. Поэтому обязательный прием — внесение органических удобрений до сева и ранневесенние подкормки. Для получения высокого урожая необходимо в первую очередь вносить в почву высокие дозы азота и калия. Фосфор и калий повышают зимостойкость растений. |


Особенности строения корневой системы щавеля и свёклы?
Песок, свешиваясь с водой, образует смесь в виде сгустка.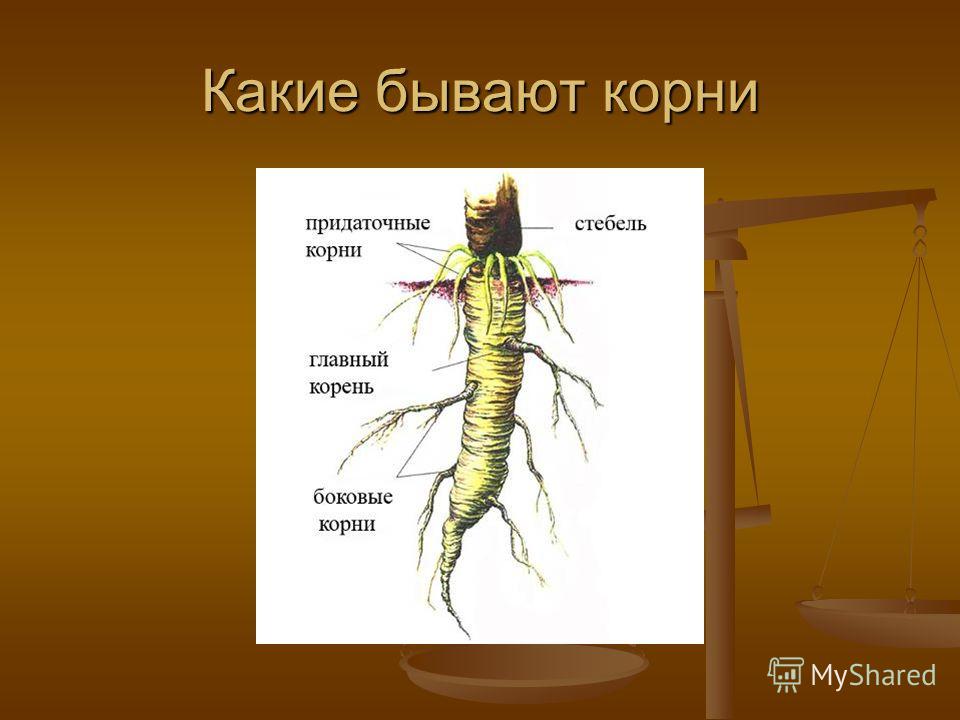 Песок- это крупинки и когда мы фильтруем смесь, то песчинки ( песок) останется в фильтре. Можно по разному пытаются отфильтровать смесь песка и воды, но этот способ самый простой и часто используемый.
Песок- это крупинки и когда мы фильтруем смесь, то песчинки ( песок) останется в фильтре. Можно по разному пытаются отфильтровать смесь песка и воды, но этот способ самый простой и часто используемый.
Каменный уголь – это осадочная порода, образовавшаяся при разложении остатков растений (древовидных папоротников, хвощей и плаунов, а также первых голосеменных растений). Основная часть добываемых в настоящее время каменных углей образовалась примерно 300-350 миллионов лет тому назад.
Дыхательная система земноводных представлена легкими и кожей, через которую они также способны дышать. Легкие – это парные полые мешки, имеющие ячеистую внутреннюю поверхность, которая усеяна капиллярами. … Потом дно ротовой полости поднимается, и ноздри снова закрываются клапанами, а воздух нагнетается в легкие.
Для пресмыкающихся характерно дыхание всасывающего типа путём расширения и сужения грудной клетки при помощи межрёберной и брюшной мускулатуры. Попавший через гортань воздух поступает в трахею — длинную дыхательную трубку, которая на конце делится на бронхи, ведущие в лёгкие.
Попавший через гортань воздух поступает в трахею — длинную дыхательную трубку, которая на конце делится на бронхи, ведущие в лёгкие.
Для дыхательной системы птиц характерно отсутствие надгортанника. У них есть нижняя гортань и воздухоносные мешки, которые являются резервуаром для воздуха, облегчают полёт и предохраняют тело птиц от перегревания. Одной из особенностей является «двойное дыхание.
1-3 2-5 3-4 4-6 5-2 6-1 я написав би так
Лук особенности строения корневой системы
Главная » Лук » Лук особенности строения корневой системыМочковатая корневая система: особенности строения и функций
Похожие статьи
Что такое корень?
Папоротниковидные
Многолетние и однолетние травы.
Типы корневых систем
Совокупность всех корней растения образует корневую систему. Корневая система формируется в течение всей жизни растения. Ее формирование обеспечивают преимущественно боковые корни. Различают два типа корневой системы: стержневую и мочковатую.
3.У фасоли, относящейся к классу Двудольные, стержневая корневая система.
Корни лука очень слабые, похожие на струны. Они мало ветвятся и имеют небольшое количество корневых волосков. Очень быстро высыхают на воздухе. Новые корни появляются в процессе роста по периферии донца. Корни лука проникают на глубину до 50 см, большая их часть находится в поверхностном слое почвы. Для хорошего роста им требуется короткий световой день и температура +5-10 градусов, а также воздух, поэтому лук очень отзывчив на рыхление. В стороны корни распространяются до 30см. Употреблять в пищу корни лука нельзя. Многолетние луки часто имеют корневище. Это не корень, а видоизмененный стебель, находящийся в почве. На корневище находятся почки, дающие начало новым побегам.
Мочковатая корневая система
Таким образом, мочковатая корневая система характерна для многих растений и обеспечивает важнейшие функции роста, питания и размножения.
Мочковатая корневая система характерна для растений, которые имеют луковицы. В них накапливается вода с растворенными питательными веществами. Тюльпан, лилия, лук-порей, чеснок используют их в качестве запаса. Это помогает им пережить неблагоприятный период.
Длина мочковатой корневой системы варьируется. У преобладающего большинства злаковых растений она достигает трех метров, а у кукурузы — до десяти. У одних из самых ценных представителей однодольных — пшеницы и ржи — основная масса придаточных корней развивается на глубине до нескольких десятков сантиметров. Поэтому такие растения очень чувствительны к недостатку влаги.
Среди двудольных подземный орган такого типа есть у подорожника. Мочковатая корневая система развивается на видоизменениях побега. Примером этого могут служить усы клубники или корневища папоротника.
Примером этого могут служить усы клубники или корневища папоротника.
Но для обеспечения всего многообразия функций подземного органа одного типа растительному организму недостаточно. Поэтому они объединяются, образуя два вида корневых систем. Стержневая состоит из главного и боковых. Ее основным преимуществом является то, что растения со стержневой системой могут получить воду глубоко из-под земли.
Растения с мочковатой корневой системой
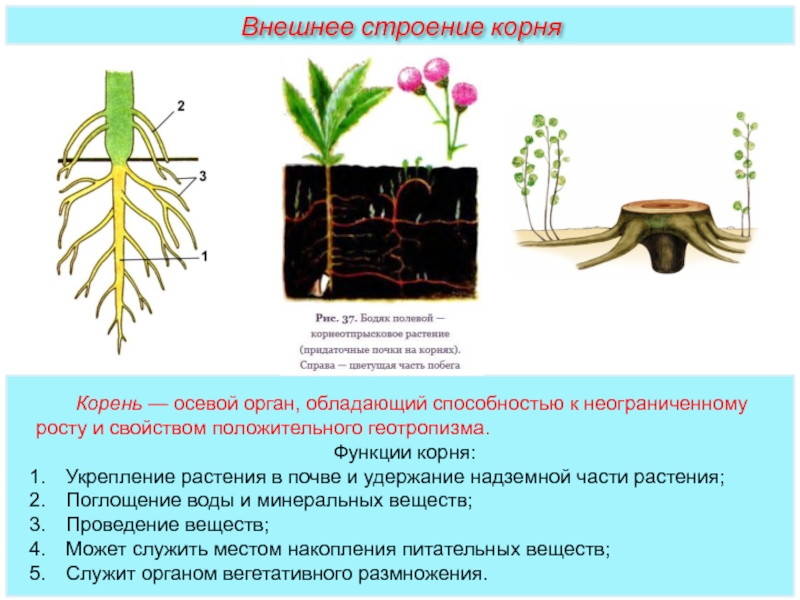
Корень выполняет в растительном организме важнейшие функции. Основные из них — закрепление в почве, всасывание и проведение воды с растворенными в ней веществами, а также вегетативное размножение. Возможность осуществления этих процессов связана с особенностями строения различных корневых систем.
Клевер ползучий
Видоизменения и их функции
————
Стержневая система имеет хорошо развитый главный корень, который развивается из зародышевого и ветвится за счет боковых корней. Главный корень способен проникать на значительную глубину.
4.Шиповник — стержневая корневая система.
fb.ru
особенности строения корневой системы лука? особенности строения корневой системы лука?
Любовь и Ирина
перр
Строение лука немного отличается от привычных нам растений. Четкого подразделения на стебель и листья, стебель и корневище у него нет. Нижние части листьев представляют собой трубчатые влагалища, из которых формируются ложные стебли. основания ложных стеблей утолщаются и образуют луковицу. В середине луковицы находится точка роста или почка.
В мочковатой корневой системе часто происходят видоизменения. В связи с этим появляются дополнительные функции. Например, придаточные корни георгина, чистяка и сладкого картофеля батата, который является востребованной культурой тропических стран, утолщаются и образуют клубни. Они не только запасают питательные вещества и воду, но и участвуют в вегетативном размножении. Воздушные корни орхидеи также являются придаточными. Они способны впитывать влагу прямо из воздуха.
Они способны впитывать влагу прямо из воздуха.
Владимир
Но у мочковатой корневой системы есть и масса преимуществ. Находясь на небольшой глубине, она охватывает гораздо большую площадь питания. Например, суммарная длина всех корней пшеницы составляет около 20 км.
Биология. 6 класс. ОСОБЕННОСТИ СТРОЕНИЯ КОРНЕВОЙ СИСТЕМЫ : фасоль, пырей, лук, шиповник, мать-и-мачеха помогите :з
kolzirin
Мочковатая корневая система способна проникать в глубь почвы на расстояние до двух метров. Там она достаточно сильно разрастается в ширину.
Мочковатая корневая система образована только придаточными корнями, которые отходят от надземной части растения — побега. Они растут крупным пучком, большинство из них имеет одинаковую длину.
Корнем называют подземный орган растения. Хотя в природе встречаются его особые разновидности — дыхательные, которые способны впитывать влагу, находящуюся в воздухе. В зависимости от особенностей строения различают несколько типов корней: главный, боковой и придаточный.
Бархатцы
Мочковатая корневая система образована совокупностью придаточных корней, растущих от стебля, и боковых корней. Главный корень в ней отсутствует. Формируется мочковатая корневая система во время кущения. На подземной части стебля при этом образуется узел кущения, из которого развиваются дополнительные побеги, то есть происходит подземное ветвление стебля, с многочисленными дополнительными корнями.
Особенности строения корневой системы у лука, пшеницы, щавеля. Пожалуйста очень надо!
Штирлиц
Такой вид корневых систем имеют многие представители двудольных растений, большинство видов деревьев и кустарников, а также многие травянистые растения.
Приведите примеры мочковатой и стержневой системы, их особенности строения корневой системы. Зарание спасибо
Ольга К.
5.Мать-и мачеха имеет мощное корневище, а на нём — многочисленные придаточные корни.
1.Корневая система лука — мочковатая (т. к. он относится к классу Однодольные). Мочка корней отрастает от видоизменённого побега — луковицы. Все корни являются придаточными.
к. он относится к классу Однодольные). Мочка корней отрастает от видоизменённого побега — луковицы. Все корни являются придаточными.
Корни лука
Мочковатая корневая система характерна и для плюща. С ее помощью он цепляется за опору и растет вверх, вынося листья к свету. Некоторые растения тропиков формируют придаточные корни прямо на стволах и ветвях. Дорастая до земли, они выполняют роль подпорок для широкой кроны. Такое же приспособление имеет и кукуруза. Поскольку придаточные корни обычно занимают поверхностное положение и не способны удержать растение в почве, эту функцию выполняют своеобразные подпорки.
Если засуха является злейшим врагом злаков и их корней, то для растений влажных природных зон она не страшна. Ведь они, наоборот, страдают от переизбытка воды. Это может вызвать процессы гниения, что неминуемо приведет к гибели растений. Именно поэтому они имеют ряд жизненно важных приспособлений для развития в этой природной зоне. Это и листья с широкой листовой пластинкой, и тонкая кора деревьев. Особое значение имеет и строение подземного органа тропических растений. Большое количество придаточных поверхностных корней обеспечивает быстрое всасывание достаточного количества влаги. По восходящему току эта вода поступает к листьям, которые обеспечивают процесс транспирации — испарения воды с поверхности пластинки.
С начала развития главный корень этой системы начинает расти. Однако вскоре он отмирает и заменяется на придаточные стеблеродные виды.
Мочковатая корневая система характерна для представителей семейств Злаковые (Мятликовые), Луковые, Лилейные. Все они относятся к классу Однодольные.
Первый у растения всегда один. От него отходят боковые корни. Обычно их достаточно много, благодаря этому увеличивается площадь всасывательной поверхности. Корни, которые растут сразу от побега, называются придаточными.
злаки, василек луговой, лютик едкий, чай луговой, клевер средний, горошек мышиный, горошек заборный, тысячелистник обыкновенный и др.
Кокосовая пальма
Двудольные — ним относятся пищевые и кормовые растения (картофель, гречиха, соя, свёкла, бахчевые и мн. др.) ; плодовые и ягодные культуры (виноград, яблоня, смородина и др.) ; масличные (подсолнечник, арахис, тунг и др.) ; большинство видов деревьев (дуб, берёза, липа и др.) ; чай, кофе, какао и сотни важнейших лекарственных растений, пряные и ароматические растения (лавр, коричное дерево и др.) ; важнейшие волокнистые растения (хлопчатник, лён, конопля, джут и др.) ; растения, дающие каучук, камеди и смолы; многие дубильные, красильные, эфиромасличные и декоративные растения.
щавель — стержневая, лук и пшеница — мочковатая
2.Корневая система пырея усложнена тем, что вместе с придаточными корнями находится корневище, но оно является видоизменённым подземным побегом. Корневая система мочковатая.
Корень — урок. Биология, Бактерии. Грибы. Растения (5–6 класс).
Функции корня
1. Корни закрепляют растение в почве и удерживают его.
2. Корни всасывают из почвы воду с растворёнными минеральными солями и обеспечивают ими все органы растения.
3. В корне могут откладываться запасы питательных веществ.
Виды корней
Различают три вида корней: главные, придаточные и боковые.
Главный корень образуется из зародышевого корешка и появляется первым из семени при его прорастании.
Придаточными называют корни, которые появляются на стеблях, листьях, клубнях, луковицах, корневищах.
Главный и придаточные корни могут ветвиться. При этом на них образуются боковые корни.
Типы корневых систем
Совокупность всех корней растения составляет его корневую систему. В зависимости от того, какие корни в неё входят, различают \(2\) типа корневых систем:
1. стержневая корневая система состоит из одного главного и множества боковых корней.
Пример:
такая корневая система имеется у одуванчика, щавеля, моркови, свёклы и др.
2. Мочковатая корневая система образована придаточными и боковыми корнями примерно одинаковых размеров. В такой корневой системе главный корень не развит или отмирает.
Пример:
такая корневая система имеется у ржи, кукурузы, лука, подорожника и др.
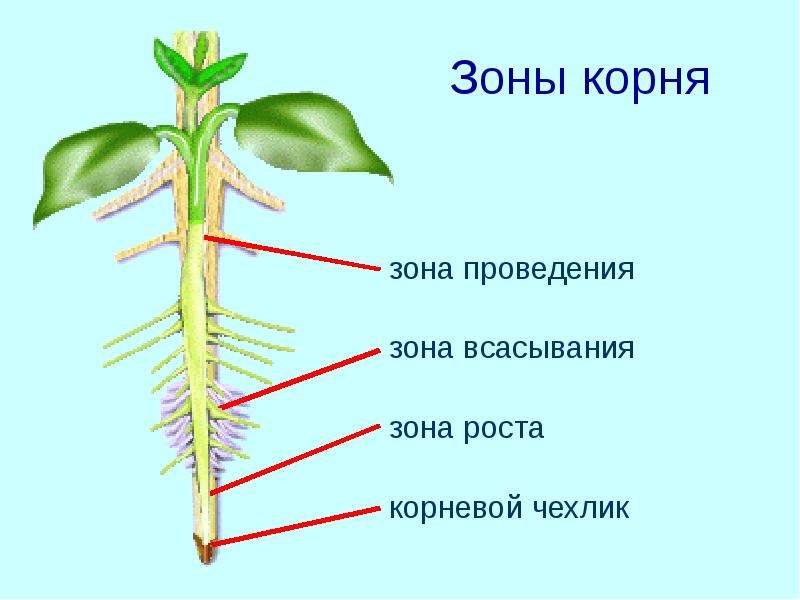
Зоны корня
На самом кончике корешка находится корневой чехлик. Он состоит из клеток покровной ткани и защищает самую уязвимую часть корня от механических повреждений. Клетки корневого чехлика живут недолго, они постоянно слущиваются и разрушаются.
Под корневым чехликом расположены клетки образовательной ткани. Это молодые мелкие клетки, которые постоянно делятся. Поэтому этот участок корня называется зоной деления.
Под чехликом расположен конус роста, состоящий из образовательной ткани. Там происходит непрерывное деление клеток. Это зона деления.
За зоной деления находится зона роста (растяжения). Здесь клетки вытягиваются, в результате чего растут в длину.
За зоной роста находится зона всасывания, в которой происходит поглощение воды и минеральных веществ корневыми волосками.
Поглощённая вода и минеральные вещества двигаются вверх по корню по зоне проведения.
Корневые волоски
Корневой волосок — это вырост клетки корня.
Большое количество корневых волосков увеличивает поверхность всасывания. Поэтому при пересадке растений корни надо беречь и перемещать с наиболее возможным количеством окружающей их почвы.
Корневые волоски под
электронным микроскопом
Корневые волоски непосредственно соприкасаются с почвой и поглощают воду и растворённые в ней минеральные вещества.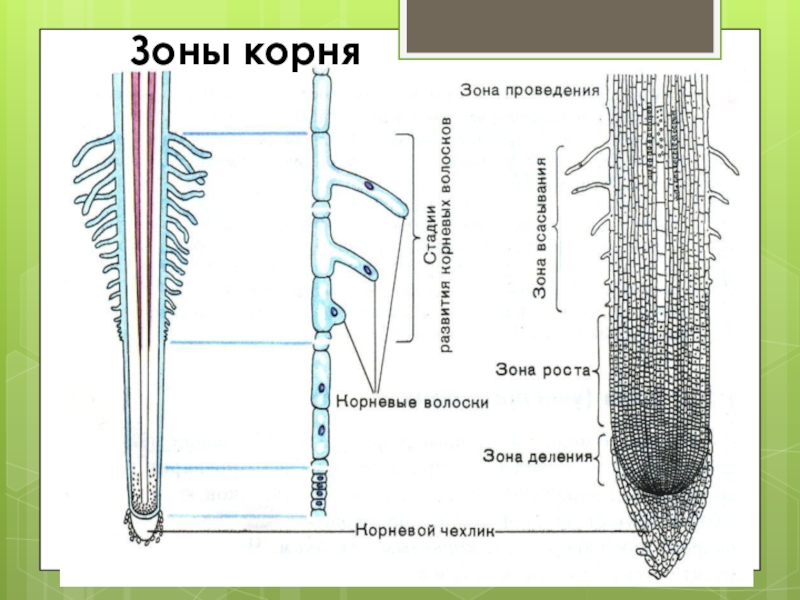
Источники:
http://all-nature.org/korni-rasteniy/
http://fullbiology.ucoz.ru/index/botanika_organy_cv_rastenij/0-293
Биометрическая классификация корневых систем деревьев
Сложное природное образование, каким является корневая система дерева, не может быть достаточно полно охарактеризовано и классифицировано только по одному какому-либо признаку.
Тип строения корневой системы должен определяться комплексом основных признаков, отражающих многообразие ее морфологических особенностей и в то же время достаточно четко выраженных. Вместе с тем это должна быть группа признаков, ограниченная по количеству, иначе формулировка типа примет расплывчатый описательный характер.
Следовательно, в группу признаков, характеризующих тип строения корневой системы или тип корневой системы, должны входить основные принципиальные особенности, от которых зависит и которыми обусловлен характер гаммы признаков, свойственных корневой системе данной древесной породы. Таким признаком является относительная интенсивность развития основных групп корней, характеризуемая их соотношением в корневой системе. В качестве основных признаков, определяющих тип строения корневой системы, целесообразно принять наличие, количественное соотношение и интенсивность развития в корневой системе трех морфологических групп корней: стержневого корня, корней горизонтальной ориентации и вертикальных ответвлений от них.
При классификации корневых систем древесных пород необходимо учитывать, что их строение изменяется с возрастом. В процессе роста в корневой системе не только количественно увеличиваются составляющие ее элементы, но и изменяется соотношение между ними. Следовательно, с возрастом может изменяться тип корневой системы. Важным для разработки и правильного понимания процесса формирования корневых систем является определение признаков, объективно отражающих возрастные особенности их строения.
Почвенно-гидрологические условия существенно влияют на структуру корневых систем, изменяя тем самым тип их строения. Это обстоятельство является серьезным препятствием в решении проблемы классификации корневых систем древесных пород.
Это обстоятельство является серьезным препятствием в решении проблемы классификации корневых систем древесных пород.
Различают следующие типы корневых систем деревьев: сформировавшиеся в соответствий с биологическими особенностями древесных пород на почвах, благоприятных для их произрастания и не оказывающих физического, химического, гидрологического и другого влияния на изменение природных особенностей строения корневых систем деревьев данной породы;
промежуточные, естественно сменяющие друг друга в возрастной динамике их формирования;
сформировавшиеся в соответствии с биологическими особенностями древесных пород, но измененные под влиянием почвенно-гидрологических и лесобиологических условий.
Тип корневой системы, представляя собой морфологическое понятие, в то же время характеризует определенные наиболее важные биологические особенности данного вида растительности. От характера морфологического строения корневой системы зависят особенности использования жизненных ресурсов, содержащихся в почвенном пространстве. Особенности пространственного размещения корневых систем предопределяют внутривидовые и межвидовые взаимоотношения растений, которые, формируясь в почвенной среде, незамедлительно отражаются на состоянии их надземной части.
В ботанике по морфологическим признакам различают два основных типа корневых систем: стержневой и мочковатый. Корневая система мочковатого типа отличается тем, что с самого начала ее образования в ней отсутствует основной (стержневой) корень или он неясно выражен. В формировании этой системы существенная роль принадлежит придаточным корням. Для корневой системы стержневого типа характерно наличие стержневого корня. Этот тип свойствен древесным и кустарниковым породам, а также определенным видам травянистой растительности (люцерна, щавель, верблюжья колючка, перекати-поле и др.).
В корневых системах стержневого типа основной корень может иметь различные характер строения, интенсивность ветвления, глубину проникновения в почву и другие морфологические признаки. Принципиально важными являются различия в соотношении количественных характеристик (массы, длины, объема, площади поверхности) стержневого корня и других морфологических групп корней. В этом отношении у древесных пород наблюдается большое разнообразие. От особенностей соотношения количественных характеристик во многом зависят интенсивность использования почвенного пространства корневой системой, формирование биологического взаимовлияния и взаимосвязи между отдельными древесными породами, а также между древесной и травянистой растительностью при совместном произрастании. В связи с этим необходимо выделение в пределах основных типов корневых систем, какими являются стержневой и мочковатый типы, более мелких их таксонов.
Принципиально важными являются различия в соотношении количественных характеристик (массы, длины, объема, площади поверхности) стержневого корня и других морфологических групп корней. В этом отношении у древесных пород наблюдается большое разнообразие. От особенностей соотношения количественных характеристик во многом зависят интенсивность использования почвенного пространства корневой системой, формирование биологического взаимовлияния и взаимосвязи между отдельными древесными породами, а также между древесной и травянистой растительностью при совместном произрастании. В связи с этим необходимо выделение в пределах основных типов корневых систем, какими являются стержневой и мочковатый типы, более мелких их таксонов.
Классификация типа строения корневой системы должна включать в себя характеристику основного общего типа ее строения, т. е. должна учитывать соответствие ее мочковатому или стержневому типу строения и характеристику таксона типа строения, т. е. определение частного типа строения. В связи с тем что все древесные породы имеют общий стержневой тип строения корневой системы, дальнейшая классификация типов корневых систем сводится к определению и упорядочению их таксонов. Таксоны типов строения корневых систем могут иметь генетическую (первая группа) и экологическую (вторая группа) обусловленность.
Генетически обусловленные типы корневых систем формируются в соответствии с наследственными, биологическими свойствами древесной породы. Это таксоны первичного порядка. Экологически обусловленные типы корневых систем формируются под влиянием конкретных экологических условий (в основном почвенно-гидрологических), существенно влияющих на таксоны первичного порядка.
Процесс формирования корневой системы в возрастной динамике представляет собой постепенную, последовательную смену типов строения корневой системы данного дерева. Например, строение корневой системы дуба, сосны или любой другой породы может существенно отличаться от строения ее в 10, 30, 55-летнем возрастах. Эти смены типов строения носят онтогенезный характер. Они обусловлены онтогенией, или онтогенезом, т. е. индивидуальным развитием организма с момента его зарождения до отмирания. Промежуточные типы строения корневых систем онтогенезного характера свойственны обеим группам таксонов.
Эти смены типов строения носят онтогенезный характер. Они обусловлены онтогенией, или онтогенезом, т. е. индивидуальным развитием организма с момента его зарождения до отмирания. Промежуточные типы строения корневых систем онтогенезного характера свойственны обеим группам таксонов.
Таким образом, классификация типов строения корневых систем представляет схему значительной сложности, отражающую многофакторность процесса их формирования и выраженную определенными морфологическими признаками.
Морфологическая характеристика корневых систем древесных пород не только имеет познавательное, теоретическое значение, но и позволяет совершенствовать лесокультурные и лесохозяйственные мероприятия в направлении повышения продуктивности лесных насаждений.
Морфология корневых систем древесных пород характеризуется их строением, т. е. количественным выражением соотношения корней различных порядков ветвления по их группам: горизонтальные, стержневые, вертикальные ответвления от горизонтальных корней. Структуру корневой системы нужно рассматривать как одну из наиболее важных характеристик, дающую возможность сравнительного анализа морфологии корневых систем отдельных деревьев и древесных пород и их классификации по типам.
Данные о структуре корневой системы дополняются биометрическими и стереометрическими характеристиками: коэффициентом ветвистости корней, коэффициентом формы корней на соответствующей относительной длине; характером связи между диаметром и суммарной длиной материнских корней первого порядка, а также между диаметром и суммарной длиной этих корней со всеми их ответвлениями; протяженностью наиболее крупных корней и их среднегодовым приростом; площадью горизонтальной проекции корневых систем; объемом почвенного пространства, занимаемого корневой системой; показателем корненаселенности указанного объема почвы, или показателем компактности корневой системы.
Кроме указанных характеристик, важное значение имеют глубина проникновения стержневого корня и вертикальных ответвлений от горизонтальных корней, а также размещение корней по почвенным горизонтам. В качестве признака, генетически присущего той или другой древесной породе, необходимо использовать соотношение массы корневой системы и других частей дерева — ствола, ветвей, ассимилирующего аппарата. Такие комплексные характеристики можно получить при полной раскопке или отмывке корневых систем от почвы с последующим анализом их в полевых и лабораторных условиях по единой методике биометрических и стереометрических исследований.
В качестве признака, генетически присущего той или другой древесной породе, необходимо использовать соотношение массы корневой системы и других частей дерева — ствола, ветвей, ассимилирующего аппарата. Такие комплексные характеристики можно получить при полной раскопке или отмывке корневых систем от почвы с последующим анализом их в полевых и лабораторных условиях по единой методике биометрических и стереометрических исследований.
В общебиологическом значении тип строения корневой системы, формировавшейся в оптимальных почвенно-гидрологических и климатических условиях, является индивидуальным для каждой древесной породы и представляется биоморфологической формулой ее корневой системы. В принципе этот тип строения представляет частный случай генетически обусловленного типа.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
§ Стержневая и мочковатая корневые системы
Изображения обложек учебников приведены на страницах данного сайта исключительно в качестве иллюстративного материала (ст. 1274 п. 1 части четвертой Гражданского кодекса Российской Федерации)
Авторы: Лабораторные работы
Вид УМК: лабораторные работы
На данной странице представлено детальное решение задания § Стержневая и мочковатая корневые системы по биологии для учеников 6 классa автор(ы) Лабораторные работы
Ход работы:
Рассматриваем корневые системы предложенных растений. Отличаются они тем, что у стержневой системы больше всего развит главный стержень, от которого уже растут тонкие корешки – придаточные. У мочковатой системы нет главного корня (он слабо развит или отмирает рано), поэтому она состоит исключительно из боковых и придаточных корней.
Отбираем растения со стержневой корневой системой. Это двудольные растения, например, свёкла, щавель, морковь.
Отбираем растения с мочковатой корневой системой. Это однодольные растения, например, лук, чеснок, пшеница, ячмень.

По строению корневой системы определяем, какие растения однодольные, какие — двудольные.
Заполняем таблицу «Строение корневых систем у разных растений».
| Пшеница | Мочковатая | Развиты придаточные и боковые корни |
| Ясень узколистый | Стержневая | Развит главный корень |
| Яблоня | Мочковатая | Развиты придаточные и боковые корни |
| Репа | Стержневая | Развит главный корень |
| Морковь | Стержневая | Развит главный корень |
| Липа | Мочковатая | Развиты придаточные и боковые корни |
| Боярышник гладкий | Стержневая | Развит главный корень |
| Лук | Мочковатая | Развиты придаточные и боковые корни |
Add
© 2021Copyright. Все права защищены. Правообладатель SIA Ksenokss.Адрес: 1069, Курземес проспект 106/45, Рига, Латвия.
Тел.: +371 29-851-888 E-mail: [email protected]
Презентация по биологии «Корень» (6 класс)
Урок биологии в 6-м классе
Тема урока: «Корень. Корневые системы».
Тип урока: урок-усвоение новых знаний.
Вид урока: урок-исследование.
Методы обучения: исследовательский, поисковый.
Формы работы: фронтальная, парная, индивидуальная, групповая.
Оборудование:
Демонстрационное –компьютер, экран, мультимедиапроектор.
Лабораторное — гербарий “Типы корневых систем”.
Цель урока: ознакомление учащихся с корнем как функциональной структурой растительного организма; формирование знания о видах корней и типах корневых систем.
Задачи урока:
1. Сформировать у школьников знания о видах корней и типах корневых систем, Познакомить с функциями корня. Изучить особенности строения корневых систем двудольных и однодольных растений.
2. Продолжить формирование умений работать с биологическими терминами, учебником, развивать практические умения по распознаванию и определению корней и корневых систем. Объяснять новые понятия, анализировать результаты своей деятельности и делать выводы. Развивать творческое мышление и монологическую речь.
3. Продолжить воспитание у учащихся коммуникативных навыков, воспитывать интерес к предмету.
Этапы урока:
Организационный момент (1 мин.)
Актуализация знаний (5 мин.)
Мотивация, сообщение цели и темы урока (2 мин.)
Изучение нового материала (20 мин.)
Обобщение и систематизация знаний (4 мин.)
Закрепление изученного материала (6 мин)
Домашнее задание (1 мин.)
Итог урока, рефлексия. (1 мин.)
ХОД УРОКА:
1. Организационный момент.
Создание доброжелательной обстановки в классе. Психологический настрой учащихся на урок
Цель: Развитие умений самооценки готовности к уроку.
2. Этап — актуализация знаний:
Проверка Д/З.
1. Рассказ с биологическими ошибками.
Мне нравятся уроки биологии. Особенно люблю работать с микроскопом. Недавно мы узнали о строении семян однодольных, двудольных и трехдольных растений. Нелегко даются знания, но я все поняла, знаю, например, что любое семя состоит из кожуры и питательных веществ. Все семена имеют эндосперм, из которого развивается новое растение. Еще я узнала, что однодольные семена у таких растений, как фасоль, тыква, а двудольные семена – у пшеницы, кукурузы.
Как все это интересно! И, главное, я хорошо во всем разобралась, как вы только что услышали!
2. Вставить пропущенные слова
(у каждого учащегося на парте лежит задание, после проверки, которого учитель обращает внимание на то, что этот текст в течение урока понадобится)
Семя состоит из …, … и содержит запас …. ….. Запас питательных веществ семени находиться в …. В зародыше различают …, …, … и …. Растения, имеющие в зародыше семени одну семядолю, называют … если же в зародыше семени две семядоли, то такие растения называют …. Горох и яблоня – это … растения. А лук и кукуруза … растения.
….. Запас питательных веществ семени находиться в …. В зародыше различают …, …, … и …. Растения, имеющие в зародыше семени одну семядолю, называют … если же в зародыше семени две семядоли, то такие растения называют …. Горох и яблоня – это … растения. А лук и кукуруза … растения.
3. Этап мотивации:
Цель: актуализация потребностно — мотивационных состояний
Учитель: Ребята, отгадайте загадку:
Царства темного жильцы
Ползут в разные концы,
Неустанно влагу пьют,
Всем растениям жизнь дают.
Учитель: Что же это? (Правильно, корни).
Учитель: Как вы думаете: о чем сегодня будет идти речь на уроке? (О корнях)
Тема нашего урока: «Корень. Корневые системы»
(Учитель читает стихотворение):
Мы в букет собрали маки жаркие,
Много незабудок голубых.
А потом цветов нам стало жалко,
Снова в землю посадили их.
Только ничего не получается:
От любого ветерка качаются!
Почему осыпались и вянут?
Без корней расти и жить не станут!
Как ни тонок, неприметен
Под землёю корешок,
Но не может жить на свете
Без него любой цветок!
(В.Жак)
Анализ стихотворения по вопросам:
Какова главная мысль стихотворения?
О каком органе цветкового растения идёт речь?
Формулировка цели урока с учащимися из стихотворения (Изучить функции корня, виды корней, типы корневых систем)
4. Изучение нового материала.
Ребята! Сегодня у нас необычный урок, а урок – исследование, в ходе которого вы будете выступать в роли исследователей.
А объектом вашего исследования будут корни и корневые системы.
Учитель: почему тему «Корень» мы изучаем после темы «Семена однодольных и двудольных растений»?
Мнение учащихся: корень развивается из зародышевого корешка, а зародышевый корешок из семени.
Учитель сообщает учащимся задачу урока:
1. Вам необходимо исследовать функции корня строение корня и изучить виды корней.
Функции корня:
Учитель: на слайде предложены картинки, по которым нужно определить функции корня.
Учитель: Корень всасывает из почвы воду и минеральные вещества.
1. Всасывание
Учитель:
Когда дует сильный ветер, попробуйте удержать за ручку зонтик. Трудно! Какая же сила нужна, чтобы дуб или липу с толстыми ветвями удержать на месте при ветре или урагане? Сила могучего великана! Этой силой обладают корни. Словно стальные канаты натянуты они во все стороны и держат растение. Корни очень крепки.
2. Укрепляет растение в почве.
3. Запасание питательных веществ
4. Вегетативное размножение
Учитель: В течение жизни у растения формируется много корней.
Исследование видов корней по группам
Учитель задает вопросы по группам (3). Учащиеся от каждой группы зарисовывают у доски виды корней
Вопрос 1:
Каким образом развивается главный корень? (Учащиеся работают с учебником.
Уч. стр. 89, 2 обзац).
Вопрос 2:
Каким образом образуются придаточные корни? (См. уч. стр. 90)
Вопрос 3:
Как возникают боковые корни? (Стр. 51)
Рассмотрели рисунок, нашли все виды корней.
Различают следующие виды корней:
Виды корней
Главный Боковые Придаточные
Учитель вместе с учащимися формулирует вывод
Совокупность всех корней растения образует единую корневую систему. В состав корневой системы входят корни разной морфологической природы: главный, боковые и придаточные.
Типы корневых систем: Стержневая, Мочковатая.
Задача 2. Вам необходимо исследовать типы корневых систем (стержневая и мочковатая) и их строение.
Учитель: Ребята как вы думаете, зная строение семени можно определить тип корневой системы?
Проблема: можно ли по строению семени определить тип корневой системы
Учитель: Для этого нам необходимо вспомнить материал предыдущего урока, где мы все растения делили на однодольные и двудольные в зависимости от строения семени.
Ребята вспоминают строение семени однодольных и двудольных растений. Приводят примеры однодольных и двудольных растений. (показывают их на слайде)
Гипотеза: зная строение семени однодольных и двудольных растений, мы сможем определить тип корневой системы.
Для этого необходимо выполнить лабораторную работу:
Лабораторная работа «Типы корневых систем. Строение стержневой и мочковатой корневых систем»
Цель: исследовать строение стрежневой и мочковатой корневых систем.
Оборудование: гербарий растений.
Задания.
Рассмотрите на гербарном материале корневые системы предложенных вам растений. Чем они различаются?
Прочитайте в учебнике, какие корневые системы называют стержневые, какие – мочковатые.
Отберите растения со стержневой корневой системой.
Отберите растения с мочковатой корневой системой.
По строению корневой системы определите, какие растения однодольные, какие – двудольные.
Заполните таблицу.
Название растения
Особенности строения корневой системы
Тип корневой системы
Однодольное или двудольное растение
1.
2.
3.
Сделайте вывод о сходствах и различиях стержневой и мочковатой корневых систем.
(мочковатую корневую систему имеют однодольные растения, стержневую корневую систему имеют двудольные растения)
Подтвердите или опровергните гипотезу.
Физминутка
Это — правая рука,
Это — левая рука.
Справа — шумная дубрава,
Слева — быстрая река…
Обернулись мы, и вот
Стало всё наоборот.
Слева — шумная дубрава,
Справа — быстрая река.
Неужели стала правой
Моя левая рука?
5. Обобщение и систематизация знаний.
Учитель читает стихотворение. Учащимся необходимо содержанием стихотворения подтвердить изученное на уроке (коллективная работа).
Учащимся необходимо содержанием стихотворения подтвердить изученное на уроке (коллективная работа).
Для чего же служит корень?
Корень нужен для того,
Что бы удержать растение,
В почве укреплять его.
А ещё он как насос
Надо нам решить вопрос:
Как он воду доставляет,
Как он соли добывает?
Знаем, корень мы один.
Есть придаточный и главный
Боковые сверх того.
И система корневая
Тоже разная бывает.
Если в сумме всех корней
Мы увидим главный корень,
То его сравним со стержнем,
Он как стержень, что тут спорить.
Если главного мы корня среди прочих не найдём,
То такую вот систему
Мочковатой назовём.
Стержневая та система
Для двудольных характерна.
Называем мы системой
Сумму всех корней растения
Для гороха и моркови,
Для бобов и для фасоли.
У пшеницы, ржи и риса,
У тюльпана и нарцисса
Главный корень не найдёшь.
К однодольным отнесёшь.
А теперь скажи, дружок,
Коль меня понять ты смог:
Так какая же система
Для пшеницы характерна?
Почему нельзя бобы
К однодольным отнести?
6. Закрепление изученного материала.
Найдите соответствие (учащимся на парту дается таблица, стрелками необходимо соединить основные понятия с правильным определением понятий).
Учащиеся меняются работами и проверяют друг у друга задание по эталону, предложенному учителем на слайде.
Осуществляется в виде небольшого тестирования по вариантам и направленного на контроль над выполнением действий исполнительного этапа и оценку усвоения знаний учащихся по теме урока. Проверяются знания и понимание изученного на уроке.
1-вариант
Основные понятия
Определение понятий
1. Корневая система
А. Система, состоящая из главного и боковых корне. В. Совокупность корней одного растения
2. Стержневая корневая система
Стержневая корневая система
Б. Корень, развивающийся из зародышевого корешка
3. Мочковатая корневая система
В. Совокупность корней одного растения Г. Система, состоящая из придаточных и боковых
4. Главный корень
Г. Система, состоящая из придаточных и боковых
5. Придаточные корни
Д. Длинные выросты наружных клеток корня, которые находятся в зоне всасывания Е. Корни, отрастающие от стеблей и листьев, касающихся земли
6. Корневые волоски
Е. Корни, отрастающие от стеблей и листьев, касающихся земли
(ответ: 1-в, 2-а, 3-г, 4-б, 5-е, 6-д).
2-вариант
Основные понятия
Определение понятий
1. Корни
А. система всех корней растений
2. Корневая система
Б. система, состоящая из главного и боковых корней
3. Главный корень
В. корни, отходящие по бокам от стебля
4. Боковые корни
Г. корень, уходящий в почву глубже всех
5. Придаточные корни
Д. подземные органы растений, всасывающие воду и минеральные соли, удерживающие растение в почве
6. Стержневая корневая система
Е. корни, отходящие по бокам от главного и придаточных корней
(ответ: 1-в, 2-а, 3-г, 4-б, 5-е, 6-д).
Дополнительное задание для учащихся имеющих высокие учебные возможности.
*Игра “Кто лишний” (индивидуальная работа).
Фасоль, горох, соя, лук.
Пшеница, морковь, рис, кукуруза.
Одуванчик, щавель, овёс, картофель.
*Закончите предложения.
1. Через корень растение получает из почвы … и ….
2. Если главный корень не развивается или не отличается от многочисленных других корней, то корневая система называется….
3. Все корни растения составляют его ….
4. На главном корне и придаточных корнях развиваются … корни.
7. Домашнее задание.
(дифференцированное)
1. Изучить параграф 19, знать основные понятия, рис. 59,60.
Изучить параграф 19, знать основные понятия, рис. 59,60.
2. Составьте синквейн с понятием “Корень.
8. Итог урока, рефлексия.
В конце урока ребятам предлагается небольшую анкету, которая позволяет осуществить самоанализ, дать качественную и количественную оценку уроку. Анкету необходимо выполнить в виде схемы, соединив необходимые пункты в правом и левом столбиках. Учащиеся оставляют анкету в рабочих тетрадях и сдают учителю.
1.На уроке я работал
2.Своей работой на уроке я
3.Урок для меня показался
4.За урок я
5.Мое настроение
6.Материал урока мне был
активно / пассивно
доволен / не доволен
коротким / длинным
не устал / устал
стало лучше / стало хуже
понятен / не понятен
полезен / бесполезен
Затем учитель собирает тетради. Каждый ученик получает индивидуальную оценку. Учитель проверяет лабораторную работу, задание и сообщает оценки на следующем уроке, объясняет ошибки и корректирует знания учащихся.
Красный щавель (Rumex acetosella) Research & Control Advice
Красный щавель (R umex acetosella ) — это разновидность многолетних широколистных сорняков, произрастающих в Евразии и Великобритании. Однако он также был завезен в большую часть северного полушария. Это особенно распространено в Калифорнии и Огайо, так как процветает в засушливых условиях, таких как пустыни и дюны.
Он также известен, как овечий щавель, красный щавель, кислый сорняк и полевой щавель.Как и многие другие широколистные сорняки (например, обыкновенный портулак), красный щавель безопасен для употребления в пищу. В некоторых регионах мира его используют в качестве гарнира или ингредиента салата. Однако он предлагает мало питательных веществ и может быть токсичным в больших количествах.
Вид — обычное явление на нарушенных землях. Он может закрепиться на обочинах дорог, в полях, садах, пахотных землях и вокруг бывших горных выработок, если почва не слишком кислая. Эти растения особенно хорошо знакомы фермерам, выращивающим чернику, поскольку они предпочитают подобные условия.
Хотя красный щавель является жизненно важным источником пищи для американской медной бабочки, садоводы по-прежнему считают его ядовитым сорняком. Его может быть сложно контролировать и удалять, потому что его ползучая структура корня корневища способна давать новые растения при разделении или расщеплении.
Если вам нужна подходящая стратегия борьбы с сорняками или ухода за газоном для красного щавеля в вашем саду, сначала убедитесь, что вы положительно определили вид. Одна из его наиболее отличительных черт — плотные участки надземных корней, которые образуются, когда растение созревает.Эти корни желтые, но могут иметь розоватый оттенок.
У растения стреловидные листья с двумя небольшими лопастями у основания. Растет близко к земле, поэтому редко поднимается на значительную высоту (максимум 0,4 м). Его стебли красного или ржаво-коричневого цвета, а цветки очень мелкие. Они растут гроздьями прямо на верхушках стеблей. Женские цветы зеленые, а мужские — желтые или красные, но оба могут существовать на одном и том же растении в одно и то же время.
Лучший способ борьбы с этим сорняком — осторожное выдергивание.Пока все растение и его корневая структура корневища удалены, оно не должно расти снова или угрожать вашему газону. Лучше всего уничтожить или изолировать корни после удаления. Всегда подумайте о том, чтобы нанять службу по уходу за газоном, чтобы позаботиться об этом сорняке.
Красный щавель
Oxalis oregana карта ареала. База данных USDA PLANTS.
Щавель красного дерева ( Oxalis oregana ). Фото доктора Джерри Карра.
Щавель красный ( Oxalis oregana ).Фото доктора Джерри Карра.
Щавель красного дерева ( Oxalis oregana ). Фото доктора Джерри Карра.
Щавель красный (
Oxalis oregana )Расс Холмс
Красный щавель относится к семейству щавелевых — Oxalidaceae, относительно небольшому семейству, состоящему из 3 родов и 875 видов в основном из тропических и субтропических регионов. Щавель красный можно использовать как почвопокровное растение в культурных ландшафтах, если условия являются тенистыми и прохладными.Растения, используемые для озеленения, можно приобрести в местных питомниках растений. Семена также можно собирать в естественной среде обитания, но только после получения надлежащего разрешения и разрешений от соответствующего агентства или землевладельца. Сбор целых растений из диких популяций категорически не рекомендуется.
Щавель красного дерева — это многолетнее растение, растущее на 5-20 см (2-7,9 дюйма) из чешуйчатого корневища. Листья все прикорневые, их обычно меньше 10. Черешки 3-20 см (1.2-7,9 дюйма) в длину и покрыты коричневыми волосками. Листовые пластинки похожи на клеверные и имеют длину от 1 до 4,5 см (0,4-1,8 дюйма). Одиночные цветы возникают выше высоты листьев на стеблях (называемых цветоносами), которые покрыты коричневыми волосками, похожими на черешки. Лепестки от белого до розового цвета и длиной 13-20 мм (0,5-0,8 дюйма). Чашелистики 5-10 мм (0,2-0,4 дюйма) в длину.
Этот вид растет в прохладных влажных лесах из ели Дугласа и прибрежных секвойи в Каскадных, Олимпийских и Прибрежных горах от юго-запада Британской Колумбии до района залива Сан-Франциско в Калифорнии.Приспособленный к тенистой среде, щавель красный способен к фотосинтезу при низкой освещенности. Свет более высокой интенсивности может повредить чувствительные листья. В качестве защитной меры листья складываются вниз в течение нескольких минут при попадании прямого света — процесс, известный как никтинастия.
Листья щавеля красного можно употреблять в пищу в вареном или сыром виде, но они умеренно токсичны из-за присутствия щавелевой кислоты. При употреблении их следует есть в небольших количествах. Известно, что северо-западные племена ели щавель красного дерева с сушеной рыбой.Коренные американцы готовили отвары из цельного растения для мытья частей тела, пораженных ревматизмом. Приготовленную из растения припарку прикладывали к фурункулам и язвам и использовали для выведения инфекции.
Для получения дополнительной информации
Rumex acetosella
Rumex acetosellaУказатель видовой информации
ВИД: Rumex acetosella
Вводный
ВИД: Rumex acetosella
АВТОР И ЦИТИРОВАНИЕ: Эссер, Лора Л.1995. Rumex acetosella. В: Информационная система по пожарным эффектам, [Интернет]. Министерство сельского хозяйства США, Лесная служба, Исследовательская станция Скалистых гор, Лаборатория пожарных наук (продюсер). Имеется в наличии: https://www.fs.fed.us/database/feis/plants/forb/rumace/all.html []. Редакций: 5 июня 2018 г. общее название этого вида было изменено в FEIS. из: овечий щавель к: щавель обыкновенный овечий. Также были добавлены изображения.
СОКРАЩЕНИЕ: РУМАС СИНОНИМЫ: Acetosella vulgaris Fourr. КОД ЗАВОДА NRCS: RUAC3 ОБЩИЕ НАЗВАНИЯ: щавель обыкновенный красный щавель овечий щавель ТАКСОНОМИЯ: Научное название щавеля обыкновенного - Rumex acetosella L. [29,34,51,95]. Он принадлежит к семейству Polygonaceae. Нет распознаются инфратаксы [34,44]. ЖИЗНЕННАЯ ФОРМА: Forb ФЕДЕРАЛЬНЫЙ ПРАВОВОЙ СТАТУС: Нет особого статуса ДРУГОЙ СТАТУС: ЗАПРЕЩЕНИЕ
РАСПРОСТРАНЕНИЕ И ПРИСУТСТВИЕ
ВИД: Rumex acetosella
ОБЩЕЕ РАСПРЕДЕЛЕНИЕ: Щавель обыкновенный - натурализованный дикий хищник евразийского происхождения. на большей части территории Северной Америки с умеренным климатом [46,75,95].
| Распределение щавеля обыкновенного. Карта любезно предоставлена Министерством сельского хозяйства США, NRCS. 2018. База данных РАСТЕНИЙ. Национальная группа данных по растениям, Гринсборо, Северная Каролина [5 июня 2018 г.] [88]. |
ЭКОСИСТЕМЫ: Щавель обыкновенный встречается в большинстве экосистем FRES. ГОСУДАРСТВА: AK AZ AR CA CO CT DE FL GA HI ID IL IN IA KS KY ME MD MA MI MN MO MT NE NV NH NJ NM NY NC ND OH OK ИЛИ PA RI SC SD TN TX UT VT VA WA WV WI WY AB BC MB NB NS ON PQ SK BLM ФИЗИОГРАФИЧЕСКИЕ ОБЛАСТИ: 1 северная граница Тихого океана 2 Каскадные горы 3 южная граница Тихого океана 4 горы Сьерра 5 Плато Колумбия 6 Верхний бассейн и хребет 8 северных скалистых гор 9 Средние Скалистые горы 10 Бассейн Вайоминга 11 Южные Скалистые горы 12 Плато Колорадо 13 Скалистые горы Пьемонт 14 Великих равнин 15 Поднятие Блэк-Хиллз 16 Верхний бассейн Миссури и Расколотые земли ОБЪЕДИНЕНИЕ ЗАВОДОВ KUCHLER: ВЪЕЗД ЗАПРЕЩЕН ТИПЫ КРЫШКИ БЕЗОПАСНОСТИ: Щавель обыкновенный встречается в большинстве укрытий SAF. SRM (RANGELAND) ТИПЫ КРЫШКИ: 101 пырея голубая 102 Овсяница Айдахо 103 Овсяница зеленая 108 Овсяница альпийская Айдахо 110 Ponderosa сосна луговая 201 Голубой дубовый лес 203 Прибрежные леса 204 Северный прибрежный кустарник 215 Долина луга 216 горных лугов 304 Idaho fescue-bluebunch пырей 409 Высокий запрет 411 Осиновый лес 601 Прерия Блюстем 804 Овсяница высокорослая 809 Смешанная древесина твердых пород и сосна ВИДЫ МЕСТО ОБИТАНИЯ И СООБЩЕСТВА РАСТЕНИЙ: Щавель обыкновенный встречается в основном на пастбищах, в разнотравных прериях и горные луговые сообщества на западе Северной Америки, но также обычны в лесных сообществах по всей Северной Америке с умеренным климатом. Щавель обыкновенный распространен в пойменных и прибрежных местообитаниях. На западе Вашингтонский щавель обыкновенный встречается на гравийных отмелях и в поймах рек преобладает ива скулерская (Salix scouleriana). Другие партнеры включают клубнику Вирджиния (Fragaria virginiana), бархатную траву (Holcus lanatus), клевер белый (Trifolium repens), щавель курчавый (Rumex crispus), и тростник (Juncus effusus) [20]. В Орегоне щавель обыкновенный встречается в Сообщество прибрежных горных лугов с преобладанием Cheatgrass (Bromus tectorum) [45].В Калифорнии щавель обыкновенный встречается в пресноводных болотах. В сообществе преобладают овсяница высокорослая (Festuca arundinacea), осока (Carex spp.) и рогоз узколистный (Typha angustifolia) [18]. Щавель обыкновенный обычно встречается на старых полях, однолетних пастбищах и горные луговые сообщества. В Коннектикуте щавель обыкновенный встречается в посельхозкультурные луга голубого стебля (Schizachyrium scoparium). Ассоциированные компании включают краснолистную (Agrostis alba) и желтую осоку (Carex pensylvanica) [69].В Нью-Джерси щавель обыкновенный овец является членом В сообществе старополевых растений преобладает золотарник канадский (Solidago canadensis) [9]. Щавель обыкновенный обычно встречается в южных Аппалачах. травянистые лысые сообщества с преобладанием горного овса (Danthonia компресса). Среди других партнеров - ежевика без шипов (Rubus canadensis), горная голубика (Vaccinium pallidum) и фиалка (Viola spp.) [56,60]. В Индиане обыкновенный овечий щавель встречается в голубых стеблях. сообщество с каркасом (Celtis occidentalis) и гладким хвощем (Equisetum laevigatum) [81].В Монтане щавель обыкновенный встречается в смешанные степные сообщества [86]. В Калифорнии щавель обыкновенный распространен на однолетних пастбищах, в горах. луговые и многолетние кустарниковые сообщества. Партнеры включают ripgut бром (Bromus rigigus), мягкие шахматы (B. hordeaceus), серебристая трава (Aira caryophyllea), мятлик Кентукки (Poa pratense), Сандберг мятлик (P. nevadensis), чертополох итальянский (Carduus pycnocephalus), дикий овес (Avena fatua) и райграс итальянский (Lolium multiflorum) [8,12,18,61]. В Национальном побережье Пойнт-Рейес, Калифорния, обыкновенный щавель встречается в сообщество прибрежных лугов с прибрежным кресс-салатом (Arabis blepharophylla), дуб ядовитый (Toxicodendron diversiloba), Калифорния барбарис (Berberis pinnata) и находящийся под угрозой исчезновения сонома колючий (Chorizanthe valida) [11,12]. Щавель обыкновенный - распространенный вид подлеска в лесных местообитаниях. по всей Северной Америке. В Пенсильвании щавель обыкновенный встречается в сосна белая восточная (Pinus strobus) - овсянка обыкновенная (D.spicata) сообщества; ассоциируются с золотарником канадским, кипреем (Epilobium angustifolium), вербейник желтый мутовчатый (Lysimachia quadrifolia), Вирджиния спрингбьюти (Claytonia virginica), лилия форели (Erythronium americanum), щавель горный (Oxalis montana) и фиалка [3,49,93]. В провинции Альберта обыкновенный овечий щавель входит в состав 80-летнего белого ель (Picea glauca) - сосна дворовая (Pinus Banksiana) - мох перьевой (Pleurozium spp.) [21]. В Айдахо щавель обыкновенный овец встречается в пихта большая (Abies grandis) / дикий имбирь (Asarum caudatum), большой пихта / пахистима (Pachistima myrsinites) и пихта большая / девятая (Physocarpus malvaceus) типы местообитаний [30,53,54,99].В Калифорнии щавель обыкновенный встречается в секвойях (Sequoia sempervirens), пихте Дугласовой. (Pseudotsuga menziesii) и дуб орегонский белый (Quercus garryana) среды обитания [22,50,83]. В Монтане и Вайоминге обыкновенный щавель встречается в альпийской тундре. среды [94].
РАССМОТРЕНИЕ УПРАВЛЕНИЯ
ВИД: Rumex acetosella
ВАЖНОСТЬ ДЛЯ ЖИВОТНОВОДСТВА И ДИКОЙ ПРИРОДА: В Аризоне щавель обыкновенный овец пасется крупный рогатый скот и овцы, но мало кормовая ценность [39].Щавель обыкновенный содержит щавелевую кислоту, которая может быть ядовитые [46,100]. В Калифорнии и Огайо обыкновенный щавель овец пасется оленями-мулами [50,70]. В Айдахо, Монтана и Висконсин едят острохвостый тетерев и рябчик семена щавеля обыкновенного [40,41,76,86]. ПРИГОДНОСТЬ: В Юте оценки вкусовых качеств щавеля обыкновенного являются удовлетворительными для крупного рогатого скота и бедные на овец и лошадей [100]. ПИТАТЕЛЬНАЯ ЦЕННОСТЬ: Уровни питательности щавеля обыкновенного соответствуют требованиям олень-мул [50].Показатели содержания белка и энергии в щавеле обыкновенном: бедные. Пищевая ценность оценивается как низкая для водоплавающих птиц и удовлетворительная для лось, олень-мул, горная дичь, мелкие неигровые птицы и мелкие млекопитающие [100]. ЗНАЧЕНИЕ КРЫШКИ: В Юте показатели покрытия щавеля обыкновенной овцы оцениваются как удовлетворительные для мелких млекопитающих. и мелких неигровых птиц и бедных для горных птиц и водоплавающих птиц [100]. ЦЕННОСТЬ РЕАБИЛИТАЦИИ НАРУШЕННЫХ МЕСТ: Щавель обыкновенный заселяет нарушенные участки, такие как сплошные вырубки, русла рек, и открытые заминированные земли.Используется для озеленения нарушенных земель, хотя он имеет низкий рейтинг для борьбы с эрозией, а также краткосрочные и долгосрочный потенциал восстановления растительного покрова [100]. В горно-металлургическом районе из Садбери, Онтарио, щавель обыкновенный, установленный при тонком опрыскивании известняка был внесен в почву [97]. В восточно-центральном Техасе восстановление восьми невосстановленных рудников произошло естественно. Эти участки и прилегающий к ним не заминированный участок были взяты образцы для определения изменений растительности с течением времени.Процент частота встречаемости щавеля обыкновенного была следующей [79]: 3 м * 6 м 5 лет 10 лет 15 лет 20 лет 30 лет 50 лет контроль 0 11 4 6 4 3 0 0 0 * m = месяцы с момента первого отбора проб, y = годы с момента первого отбора В Пенсильвании была предпринята попытка пересадки подвоев овец. щавель на полигонах черных отходов, образовавшихся при добыче антрацита. Все возникающие впоследствии растительность была уничтожена жарой [77]. ИСПОЛЬЗОВАНИЕ И ЗНАЧЕНИЯ: Листья щавеля обыкновенного используются в салатах [4,51]. Индейцы нуксалк Британская Колумбия ест щавель обыкновенного барана [55]. ДРУГИЕ СООБРАЖЕНИЯ РУКОВОДСТВА: Щавель обыкновенный классифицируется как ядовитый сорняк в 23 штатах [67]. Это серьезный сорняк на пастбищах и пастбищах. Сложно контролировать, потому что многолетних ползучих корневищ [4,52]. Щавель обыкновенный - обыкновенный сорняк в Западной Вирджинии, за исключением известняковых регионов; известкование почвы может помочь искоренить щавель обыкновенного барана [82]. Присутствие и обилие щавеля обыкновенного указывает на плохую и «кислую» пищу. почвы [82,87]. Достигает максимальной численности при низком уровне азота в почве. [87]. Щавель обыкновенный потенциально ядовит для домашнего скота из-за наличие растворимых оксалатов [19]; однако его пасут овцы и крупный рогатый скот [39]. В Айдахо обыкновенный овечий щавель - вид-увеличитель при тяжелых условиях. режимы выпаса и уменьшающиеся виды при легких режимах выпаса [54]. В Орегоне процентная частота выращивания щавеля обыкновенного не зависела от поздний сезон выпаса крупного рогатого скота на прибрежных горных лугах [45].В Новой Шотландии щавель обыкновенный - один из самых распространенных видов сорных растений. низкорослые черничные поля. Была предпринята попытка контроля с помощью гексазинона, но после снижения активности гербицида щавель обыкновенный вырос и дали большое количество семян [62]. В Пенсильвании в золотарнике (Solidago spp.) - сообщество астры (Aster spp.), Преобладал щавель обыкновенный. на пахоте, дисковых дисках, обработанных прометоном делянках в возрасте 1 и 3 года [64]. Методы борьбы: постепенно повторяйте культивирование в засушливую погоду. ослабляет корневища щавеля обыкновенного [19].Согласно Фитцсиммонсу [19] Некоторые гербициды могут избирательно контролировать щавель обыкновенный.
БОТАНИКО-ЭКОЛОГИЧЕСКИЕ ХАРАКТЕРИСТИКИ
ВИД: Rumex acetosella
ОБЩИЕ БОТАНИЧЕСКИЕ ХАРАКТЕРИСТИКИ: Щавель обыкновенный - это интродуцированное корневищное многолетнее растение, которое иногда образует плотные колонии из придаточных побегов от широко раскиданных корней и корневища [19,29,35,52,92]. Стебли прямостоячие, тонкие, от 4 до 24. дюймов (10-60 см) в высоту [33,72,95].Листья от 0,8 до 4 дюймов (2-10 см). в длину и от 0,4 до 0,8 дюйма (1-2 см) в ширину [33,34,72]. Плод - это семянки [25,29,38,72,95]. Корни тонкие, почти волокнистые, проникать на глубину 5 футов (1,5 м) [48]. ЖИЗНЕННАЯ ФОРМА РАУНКИАРА: Гемикриптофит Геофит ПРОЦЕССЫ РЕГЕНЕРАЦИИ: Половой признак: Щавель обыкновенный размножается семенами. Опыляется ветром и семена разносятся ветром и насекомыми [37,91].Щавель обыкновенный регулярно колонизируется из заглубленных семян после нарушения [14,21,26]. В Щавель обыкновенной овцы Массачусетса не присутствовал в почвенном покрове большинства восточная белая сосна и красная сосна (Pinus Resinosa) насаждения, но семена были содержится в образцах почвы от 1 до 80-летних насаждений. в семена лабораторного хранения в почве из всех насаждений проросли [57]. Вегетативный: щавель обыкновенный размножается из ползучих корней и корневищ. [2,16,48,77].Побеги развиваются из стволовых почек, возникающих случайно. через нерегулярные интервалы на горизонтальных корнях. Придаточные бутоны обычно встречается в верхних 8 дюймах (20 см) почвы [48]. ХАРАКТЕРИСТИКИ САЙТА: Щавель обыкновенный распространен на полях, пастбищах, лугах, пустырях и вдоль обочин [4,29,39,46,72,95]. В Олимпийском национальном парке, Вашингтон, щавель обыкновенный обычно встречается на нижних террасах и на гравии. бары прибрежной зоны [1,13,63].Щавель обыкновенный обычно встречается в открытые, незатененные участки на нарушенных участках [29,92,95]. Он процветает на кислые почвы с низким плодородием, но адаптированы к различным почвам типы [19,29,96]. Щавель обыкновенный обычно встречается на супеси, мелкой песчаные, илистые и гравийные почвы [6,7,14,58,99]. Отметки для щавеля обыкновенного следующие: фут-метры Аризона 5 500–8 000 1 650–2400 [46] Калифорния 0–9 900 0–3 000 [34] Колорадо 4,000-11,200 1,200-3,360 [33,100] Коннектикут 1020–1050 310–320 [14] Айдахо 2 800–6 000 853–1830 [65,99] Монтана 3 300–6 200 990–1860 [100] Нью-Йорк 900–1700 270–510 [85] Орегон 66] Юта 4 500–9 100 1 350–2730 [95 100] Вашингтон 66] Вайоминг 6000–9000 1800–2700 [100] ПОСЛЕДНИЙ СТАТУС: Щавель обыкновенный вторгается в нарушенные участки и может переходить на нетронутые участки. при идеальных условиях выращивания [1,16,52,77].Обычно встречается на сплошные, выжженные и нарушенные наводнением прибрежные участки [1,13,27,94]. Это быстро колонизируется семенами и может сохраняться от 15 до 20 лет через вегетативный рост и размножение [16]. Конкуренция со стороны других видов на хороших почвах может снизить его численность [19]. В Южной Каролине щавель обыкновенный встречается на ранних стадиях семенного цикла. нарушенный стебель обыкновенной метлы (Andropogon virginicus) сообщество [101]. На Тихоокеанском Северо-Западе семена щавеля обыкновенной овцы обычно сохраняют всхожесть почва достаточно длинная, чтобы стать источником новых заражений, когда почва нарушена [19].В Массачусетсе закопали семя щавеля обыкновенной овцы проросли из образцов почвы из насаждений белой и красной сосны восточной От 1 до 80 лет [57]. Щавель обыкновенный умеренно теневынослив. В предгорьях Сьерра-Невада и Прибрежные хребты, Калифорния, щавель обыкновенный овец был более больше под мертвым голубым дубом (Quercus douglasii) (5,7%), чем в открытые луга (5,2%) или живые насаждения из голубого дуба (36). СЕЗОННОЕ РАЗВИТИЕ: Сроки цветения щавеля обыкновенного следующие: Калифорния, март - август [68] Айдахо, май – сентябрь [19] Джорджия Мар-июн [98] Канзас, апрель-июль [4] Монтана, май – август [100] Северная Каролина, мар-июль [72,98] Северная Дакота, май-июнь [100] Орегон, май – сентябрь [19] Южная Каролина, мар-июль [72,98] Теннесси Мар-Джун [98] Вирджиния Мар-Джун [98] Вашингтон, май-сентябрь [19] Западная Вирджиния, май-сентябрь [82] Великие равнины, апрель – август [29]
ПОЖАРНАЯ ЭКОЛОГИЯ
ВИД: Rumex acetosella
ПОЖАРНАЯ ЭКОЛОГИЯ ИЛИ АДАПТАЦИИ: Щавель обыкновенный, вероятно, выживает при пожаре, прорастая из корневищ и корни [47,72].Вероятно, он регенерируется из семян, закопанных на месте. ПОЖАРНЫЕ РЕЖИМЫ: Найдите информацию о пожарном режиме для растительных сообществ, в которых виды могут встречаться при вводе названия вида на домашней странице FEIS в разделе «Найдите режимы огня». СТРАТЕГИЯ РЕГЕНЕРАЦИИ ПОСЛЕ ПОЖАРА: Корневищная трава, корневище в почве Наземный остаточный колонизатор (на месте, начальное сообщество) Вторичный колонизатор - посевной материал за пределами участка
ПОЖАРНОЕ ВОЗДЕЙСТВИЕ
ВИД: Rumex acetosella
НЕМЕДЛЕННОЕ ПОЖАРНОЕ ВОЗДЕЙСТВИЕ НА ЗАВОД: Обыкновенный щавель, вероятно, погиб от пожара. ОТВЕТЫ НА ПОЖАР ЗАВОДА: Щавель обыкновенный, вероятно, вырастает из корневищ после пожара и устанавливается из семян на месте [14,21,26]. В нескольких исследованиях описывается укоренение или выращивание щавеля обыкновенного после пожара. Очень сильный пожар может убить щавель обыкновенного барана. Осенью 1949 г. в Нью-Брансуике был сплошной лесной выруб. предписано сжигать в апреле 1951 г. Количество стеблей щавеля обыкновенного. на площадь, присутствующую в июне 1949, 1950, 1951 и 1952 годов, были 0, 0, 18 и 28 соответственно [31].В нижнем ярусе Нью-Брансуика 11 смешанных слоев древостои лиственных пород, представляющие возрастную последовательность от 7 до 57 лет после пожара были обследованы. Отбор проб проводился в июле и августе 1973 и 1974 гг. Щавель обыкновенный встречен в насаждениях возрастом 7, 10, 13, 17 и 25 лет. Это не встречались в некоторых 7-летних трибунах или в клетях 18, 20, 29 и 37 лет [58]. В Айдахо отдельные заросли кустарников в среде обитания большой пихты / пахистимы были предписано сожжено 14 мая 1975 г., а часть засеяна 15 мая, 1975 г.Общие щавель овец присутствовал на ожоговой только области, но не произошло на ожогово-семенном участке. Частота (из 10 возможных участков) Щавель обыкновенный овечий был следующим [53]: Prefire Постпожарный год 3 июля 1974 г. 1 2 4 контроль 1 2 3 3 только гореть 0 0 5 4 сжечь и посеять 0 0 0 0 В Айдахо лесной пожар сжег сосновый лес пондероза (Pinus ponderosa) и прилегающие горные луга 10 августа 1973 г. в течение 43 дней.Четырнадцать участки были обследованы в июне 1974 г. и июне 1976 г. Процентное покрытие и Частота появления щавеля обыкновенного на сожженных и несгоревших участках составила [65]: 1974 1976 сгорел несожженный сожженный несгоревший крышка +/- SD t * +/- 1 t +/- t 2 +/- 3 1 +/- 2 частота +/- SD 1 +/- t t +/- 1 2 +/- 4 1 +/- 2 * t = след В Вашингтоне на горе Адамс черника (Vaccinium spp.) Поля, с 3 по 7 октября 1972 г. был предписан сожженный опытный участок. Среднее покрытие подлеска (%) щавеля обыкновенного с 1972 по 1977 гг. Было примерно таким же. следует [66]: 1972 г. (до лечения) 1973 г. 1974 г. 1975 г. 1977 г. необожженные необрезанные 0,2 0,2 0,3 0,3 0,2 тонкий, недожженный 0,2 0,6 0,9 1,2 1,5 сплошные и сожженные 0,2 0,9 1,9 1,0 1,6 В Великобритании сильные пожары в конце лета 1976 года уничтожили всю поверхность. растительность.Щавель обыкновенный впервые появился на гарях в октябре 1985, год после пожара 9 [59]. ОБСУЖДЕНИЕ И КВАЛИФИКАЦИЯ ОТВЕТОВ ЗАВОДА: Некоторые исследования показывают, что у щавеля обыкновенного нет изменений в покрытии или частоте. после пожара. В Калифорнии последствия позднего осеннего ожога на горе луг в государственном парке Гровер Хот Спрингс. И мокрый, и на сухих луговых участках предписывались горения от слабой до умеренной интенсивности. Пожар в середине ноября 1987 г.Щавель обыкновенный встречен только на сухих участках. до горения и не увеличивалось после пожара [8]. В Коннектикуте экспериментальные трактаты были созданы в маленьком голубом доме. пастбищ в 1967 году. Урочище А было предписано сжигать ежегодно из 1968–1976, а также в 1978, 1980, 1983 и 1985 годах. Был назначен тракт B горели ежегодно с 1968-1975, а также в 1978, 1980, 1983 и 1985 годах. Процент покрытия щавеля обыкновенного и частота в двух ожогах и 2 контрольных участки на каждом тракте были следующие [69]: Урочище А Урочище Б 1967 1985 1967 1985 cover freq cover freq cover freq cover freq лечение Ожог
в лесах западной Монтаны
и осенних пожаров в штате Массачусетс
ССЫЛКИ
ВИД: Rumex acetosella
ССЫЛКИ: 1.Эйджи, Джеймс К. 1988. Сукцессионная динамика в прибрежных лесных зонах. В: Редеке, Кеннет Дж., Изд. Управление рекой: прибрежная дикая природа и лесохозяйственные взаимодействия. Институт лесных ресурсов Вклад No. 58. Сиэтл, Вашингтон: Вашингтонский университет, Колледж лесных ресурсов: 31-43. [7657] 2. Antos, Joseph A .; Зобель, Дональд Б. 1985. Форма растения, развитие пластичность и выживаемость после захоронения вулканической тефрой.Канадский Журнал ботаники. 63: 2083-2090. [12553] 3. Auchmoody, L.R .; Уолтерс, Р. С. 1988. Восстановление убитого рассолом лесной участок. Журнал Американского общества почвоведов. 52: 277-280. [11374] 4. Бэр, Джанет Э. 1979. Полевые цветы и сорняки Канзаса. Лоуренс, KS: Regents Press Канзаса. 509 с. [3801] 5. Бернард, Стивен Р .; Браун, Кеннет Ф. 1977. Распространение млекопитающих. рептилий и земноводных по физико-географическим регионам БЛМ и A.В. Кухлера ассоциации одиннадцати западных штатов. Tech. Примечание 301. Денвер, Колорадо: Министерство внутренних дел США, Бюро землепользования. 169 с. [434] 6. Бисвелл, Х. Х. 1956. Экология лугов Калифорнии. Журнал Лесное хозяйство. 9: 19-24. [11182] 7. Блюетт, Томас. 1978. Восстановление прерий и саванн в Неседе. Национальный заповедник дикой природы. В: Glenn-Lewin, David C .; Ландерс, Роджер К., Младший, ред. Proceedings, 5-я конференция по прериям Среднего Запада; 1976 Август 22-24; Эймс, ИА. Эймс, ИА: Государственный университет Айовы: 154-157. [3370] 8. Бойд, Роберт С .; Вудворд, Рой А.; Уолтер, Гэри. 1993. Воздействие пожара на горный луг Сьерра-Невада. Калифорнийская рыба и дичь. 70 (3): 115-125. [24152] 9. Carson, Walter P .; Пикетт, С. Т. 1990. Роль ресурсов и нарушение организации старополевого растительного сообщества. Экология.71 (1): 226-238. [11078] 10. Клампитт, Кристофер А. 1993. Влияние антропогенных нарушений на прерии. и региональный эндемичный Aster curtus в западном Вашингтоне. Северо-Запад Наука. 67 (3): 163-169. [23038] 11. Clark, Ronilee A .; Fellers, Гэри М. 1986. Редкие растения Point Reyes. Национальное побережье. Tech. Представитель № 22. Дэвис, Калифорния: Университет г. Калифорния, Институт экологии; Сан-Франциско, Калифорния: Департамент США внутренних дел, Служба национальных парков, Западный регион.117 с. [18095] 12. Дэвис, Лиам Х .; Шерман, Роберт Дж. 1992. Экологическое исследование редких Chorizanthe valida (Polygonaceae) на национальном побережье Пойнт-Рейес, Калифорния. Мадроно. 39 (4): 271-280. [19787] 13. DeFerrari, Collette M .; Найман, Роберт Дж. 1994. Многоуровневая оценка. появления экзотических растений на Олимпийском полуострове в Вашингтоне. Журнал науки о растительности. 5: 247-258. [23698] 14. Дель Тредичи, Питер.1977 г. Захороненные семена Comptonia peregrina, сладкий папоротник. Бюллетень Ботанического клуба Торри. 104 (3): 270-275. [21893] 15. Дорн, Роберт Д. 1988. Сосудистые растения Вайоминга. Шайенн, Вайоминг: гора Вест Паблишинг. 340 с. [6129] 16. Эскарре, Жозеп; Хуссар, Клоди; Томпсон, Джон Д. 1994. Экспериментальный изучение роли плотности проростков и соседства в устойчивость Rumex acetosella в последовательности старых полей.Канадский Журнал ботаники. 72 (9): 1273-1281. [24362] 17. Эйр, Ф. Х., изд. 1980. Типы лесного покрова США и Канада. Вашингтон, округ Колумбия: Общество американских лесоводов. 148 с. [905] 18. Фидлер, Пегги Ли; Лейди, Роберт А. 1987. Растительные сообщества Ринга. Горный заповедник, округ Марин, Калифорния. Мадроно. 34 (3): 173-192. [4068] 19. Fitzsimmons, J. P .; Беррилл, Л.С. 1993. Красный щавель: Rumex acetosella L.Сорняки. Корваллис, Орегон: издание Pacific Northwest Extension Publication; PNW 446: [25189] 20. Фонда Р. У. 1974. Сукцессия леса по отношению к речной террасе. развитие в Олимпийском национальном парке, Вашингтон. Экология. 55 (5): 927-942. [6746] 21. Файлз, Джеймс В. 1989. Популяции банка семян в горных хвойных лесах. в центре Альберты. Канадский журнал ботаники. 67: 274-278. [6388] 22. Гарднер, Роберт А.1958. Почвенно-растительные ассоциации секвойи - Зона Дугласовой пихты Калифорнии. В: Proceedings, 1st North American конференция по лесным почвам; [Дата конференции неизвестна]; Ист-Лансинг, штат Мичиган. Ист-Лансинг, Мичиган: Университет штата Мичиган, Сельскохозяйственный эксперимент Станция: 86-101. [12581] 23. Гаррисон, Джордж А .; Bjugstad, Ardell J .; Дункан, Дон А .; [и другие]. 1977. Растительность и экологические особенности леса и ареала. экосистемы.Agric. Handb. 475. Вашингтон, округ Колумбия: Департамент США Сельское хозяйство, лесная служба. 68 с. [998] 24. Gashwiler, Джей С. 1970. Изменения растений и млекопитающих на сплошной рубке в западно-центральный Орегон. Экология. 51 (6): 1018-1026. [8523] 25. Глисон, Генри А .; Кронквист, Артур. 1991. Определитель сосудистых растений г. северо-восток США и прилегающая Канада. 2-е изд. Нью-Йорк: Нью-Йорк Йоркский ботанический сад. 910 с. [20329] 26. Гранстрем, А.; Шиммель, Дж. 1993. Тепловое воздействие на семена и корневища растений. выбор растений бореальных лесов и потенциальная реакция на огонь. Oecologia. 94: 307-313. [22867] 27. Гранстрем, Андерс. 1982. Банк семян в пяти бореальных насаждениях. возник между 1810 и 1963 годами. Канадский ботанический журнал. 60: 1815-1821. [5940] 28. Grant, S.A .; Torvell, L .; Smith, H.K .; [и другие]. 1987. Сравнительный исследования выбора рациона овец и крупного рогатого скота: покрывное болото и вереск причалить.Журнал экологии. 75: 947-960. [21037] 29. Ассоциация флоры Великих равнин. 1986. Флора Великих равнин. Лоуренс, KS: Университетское издательство Канзаса. 1392 с. [1603] 30. Грин, Пэт; Дженсен, Марк. 1991. Правопреемство заводов в рамках управляемых грандиозных еловые леса северного Айдахо. В: Harvey, Alan E .; Нойеншвандер, Леон Ф., составители. Ход работы - управление и продуктивность западно-горные лесные почвы; 10-12 апреля 1990 г .; Бойсе, ID.Gen. Tech. Реп. INT-280. Огден, штат Юта: Министерство сельского хозяйства США, Лесная служба, Межгорная исследовательская станция: 232-236. [15987] 31. Холл, И. В. 1955. Флористические изменения после срезки и сжигания лесной участок для выращивания черники. Канадский сельскохозяйственный журнал Наука. 35: 143-152. [9012] 32. Холл, Иван В .; Aalders, Lewis E .; Никерсон, Нэнси Л .; Вандер Клоет, Сэм П. 1979. Биологическая флора Канады.I. Vaccinium angustifolium Айт., Сладкая низкорослая черника. Канадский естествоиспытатель. 93 (4): 415-430. [9185] 33. Харрингтон, Х. Д. 1964. Справочник растений Колорадо. 2-е изд. Чикаго: The Swallow Press Inc., 666 с. [6851] 34. Хикман, Джеймс С., изд. 1993. Учебник Джепсона: Высшие растения Калифорния. Беркли, Калифорния: Калифорнийский университет Press. 1400 с. [21992] 35. Хичкок, К. Лео; Кронквист, Артур.1973. Флора Тихого океана. Северо-Запад. Сиэтл, Вашингтон: Вашингтонский университет Press. 730 с. [1168] 36. Голландия, В. Л. 1980. Влияние голубого дуба на кормопроизводство пастбищных угодий. в центральной Калифорнии. В: Пламб, Тимоти Р., технический координатор. Материалы симпозиума по экологии, менеджменту и утилизации. Калифорнийских дубов; 1979 26-28 июня; Клермонт, Калифорния. Gen. Tech. Rep. PSW-44. Беркли, Калифорния: Министерство сельского хозяйства США, Лесная служба, Тихоокеанская Юго-Западная экспериментальная станция лесов и полигонов: 314-318.[7052] 37. Houssard, C .; Escarre, J. 1991. Влияние веса семян на рост и конкурентоспособность Rumex acetosella из двух последовательных старые поля. Oecologia. 86 (2): 236-242. [25190] 38. Халтен, Эрик. 1968. Флора Аляски и прилегающих территорий. Стэнфорд, Калифорния: Издательство Стэнфордского университета. 1008 с. [13403] 39. Хамфри, Роберт Р. 1955. Производство кормов на пастбищах Аризоны, IV. Коконино, Навахо, округа Апачи: исследование в условиях ареала.Бюллетень 266. Тусон, Аризона: Университет Аризоны, Экспериментальная сельскохозяйственная станция. 84 с. [5087] 40. Хангерфорд, К. Э. 1951. Популяции рябчиков и укрытие в северный Айдахо. Сделки, 16-я Североамериканская конференция по дикой природе. [Объем неизвестен]: 216-224. [13618] 41. Хангерфорд, Кеннет Э. 1957. Оценка кормов для рябчиков с точки зрения среды обитания. улучшение. Транзакции, 22-я Североамериканская конференция по дикой природе.[Объем неизвестен]: 380-395. [15905] 42. Jobidon, R .; Thibault, J. R .; Фортин, Дж. А. 1989. Фитотоксическое действие мульчи из ячменя, овса и пшеничной соломы в лесах восточного Квебека плантации 1. Воздействие на красную малину (Rubus idaeus). Экология леса и менеджмент. 29: 277-294. [9899] 43. Джудей, Гленн Патрик. 1992. Исследования природных территорий Аляски. 3: змеевик Горка. Gen. Tech. Представитель PNW-GTR-271. Портленд, Орегон: Департамент США Сельское хозяйство, лесная служба, Тихоокеанская Северо-Западная научно-исследовательская станция.66 с. [21511] 44. Кауфман, Дж. Бун; Krueger, W. C .; Вавра, М. 1983. Последствия последнего сезонный выпас скота на прибрежных растительных сообществах. Журнал диапазона Управление. 36 (6): 685-691. [16965] 46. Kearney, Thomas H .; Пиблз, Роберт Х .; Хауэлл, Джон Томас; МакКлинток, Элизабет. 1960. Флора Аризоны. 2-е изд. Беркли, Калифорния: Университет California Press. 1085 с. [6563] 47. Кеун, Ларри Д. 1978. Управление пожарами в Селуэй-Биттеррут. Пустыня, Район рейнджеров Лос-Крик, Национальный лес Нез-Персе.Миссула, MT: Министерство сельского хозяйства США, Лесная служба, Северная Область, край. 163 с. [18634] 48. Кильц Б.Ф. 1930. Многолетние сорняки, распространяющиеся вегетативно. Журнал Американское общество агрономии. 22 (3): 216-234. [25191] 49. Kolb, T. E .; Bowersox, T. W .; Маккормик, Л. Х. 1990. Влияние света. интенсивность стресса сеянцев деревьев, вызванного сорняками. Канадский журнал Лесные исследования. 20: 503-507. [12251] 50.Крюгер, Уильям С .; Донарт, Гэри Б. 1974. Связь почв с сезонное качество кормов для оленей. Журнал Range Management. 27 (2): 114-117. [25192] 51. Кудиш, Михаил. 1992. Растительный мир Адирондакского нагорья: экологическая перспектива. Саранак, Нью-Йорк: The Chauncy Press. 320 с. [19376] 52. Лакшевиц, Клаус. 1991. Сосудистые растения западно-центральных регионов. Монтана - справочник по идентификации. Gen. Tech. Реп. INT-227. Огден, штат Юта: U.S. Департамент сельского хозяйства, Лесная служба, Межгорные исследования Станция. 648 с. [13798] 53. Leege, Thomas A .; Годболт, Грант. 1985. Настоящий ответ после предписано сжигание и посев лосейного пастбища в Айдахо. Северо-западная наука. 59 (2): 134-143. [1436] 54. Leege, Thomas A .; Герман, Дэрил Дж .; Замора, Бенджамин. 1981. Эффекты выпас скота на горных лугах в Айдахо. Журнал диапазона Управление. 34 (4): 324-328.[2961] 55. Лепофски, Дана; Тернер, Нэнси Дж .; Kuhnlein, Харриет В. 1985. Определение доступность традиционных продуктов из диких растений: пример Nuxalk еда, Белла Кула, Британская Колумбия. Экология питания и питания. 16: 223-241. [7002] 56. Линдси, Мэри М .; Браттон, Сьюзан Пауэр. 1979. Травянистые лысины Великого. Смоки-Маунтинс: их история и флора по отношению к потенциалу управление. Управление окружением.3 (5): 417-430. [23347] 57. Ливингстон, Р. Б .; Алессио, Мэри Л. 1968. Захоронены жизнеспособные семена в сукцессионные поля и лесные насаждения, Гарвардский лес, Массачусетс. Бюллетень Ботанического клуба Торри. 95 (1): 58-69. [3377] 58. MacLean, David A .; Wein, Ross W. 1977. Изменения в подлеске растительности. с увеличением возраста древостоя в лесах Нью-Брансуика: видовой состав, покров, биомасса и питательные вещества. Канадский журнал ботаники.55: 2818-2831. [10106] 59. Maltby, E .; Legg, C.J .; Проктор, М. С. Ф. 1990. Экология суровых Пожар вересковой пустоши на болотах Северного Йорка: последствия пожаров 1976 года и последующее развитие поверхности и растительности. Журнал экологии. 78 (2): 490-518. [19852] 60. Марк А. Ф. 1958. Экология травяных лысин южных Аппалачей. Экологические монографии. 28 (4): 293-336. [23350] 61. Макбрайд, Джо Р.; Норберг, Эд; Ченг, Шеучи; Мосаддык, Ахмад. 1991 г. Посадка рассады прибрежного живого дуба в связи с хранением семян Джей. В: Стэндифорд, Ричард Б., технический координатор. Труды симпозиум по управлению дубовыми лесами и пастбищами лиственных пород; 1990 г. 31 октября - 2 ноября; Дэвис, Калифорния. Gen. Tech. Реп. PSW-126. Беркли, Калифорния: Министерство сельского хозяйства США, Лесная служба, Тихоокеанский юго-запад Исследовательская станция: 143-148.[19055] 62. McCully, Kevin V .; Сэмпсон, М. Глен; Уотсон, Алан К. 1991. Опрос сорняков. из Новой Шотландии (Канада) голубика низкорослая (Vaccinium angustifolium) поля. Наука о сорняках. 39 (2): 180-185. [20036] 63. Макки, Артур; ЛаРой, Джордж; Франклин, Джерри Ф. 1979. Структура, состав и репродуктивное поведение террасных лесов, Саут-Форк Река Хох, Олимпийский национальный парк. В: Материалы 2-й конференции научные исследования в национальных парках; [Дата конференции неизвестна]; [Место проведения конференции неизвестно].[Место публикации неизвестно]. [Издатель неизвестен]. 22-29. [21571] 64. Медве, Ричард Дж. 1984. Микоризы пионерных видов в нарушенных экосистемы западной Пенсильвании. Американский журнал ботаники. 71 (6): 787-794. [8544] 65. Merrill, Evelyn H .; Мэйленд, Генри Ф .; Пик, Джеймс М. 1980. Эффекты лесной пожар на травянистой растительности на ксерических участках в Селуэй-Биттеррут Уайлдернесс, штат Айдахо. Журнал Range Management.33 (5): 363-367. [1642] 66. Миноре, Дон; Смарт, Алан У .; Дубрасич, Майкл Э. 1979. Гекльберри экологические и управленческие исследования на Тихоокеанском Северо-Западе. Gen. Tech. Представитель PNW-93. Портленд, Орегон: Министерство сельского хозяйства США, лес Служба, Тихоокеанская Северо-Западная экспериментальная станция лесов и полигонов. 50 шт. [6336] 67. Mitich, Larry W .; Кайзер, Гай Б. 1992. Влияние экзотических сорняков на Соединенные Штаты. В: Лим, Родни Г., изд. Труды, Западное общество Наука о сорняках; 1992 10-12 марта; Солт-Лейк-Сити, штат Юта. [Место публикация неизвестна]. Западное общество по изучению сорняков: 86-93. [20616] 68. Munz, Philip A. 1973. Флора Калифорнии и приложение. Беркли, Калифорния: Калифорнийский университет Press. 1905 г. с. [6155] 69. Niering, William A .; Дрейер, Гленн Д. 1989. Эффекты предписанного горение Andropogon scoparius на постсельскохозяйственных лугах в Коннектикут.Американский натуралист из Мидленда. 122: 88-102. [8768] 70. Nixon, Charles M .; McClain, Milford W .; Рассел, Кеннет Р. 1970. Олень пищевые привычки и характеристики диапазона в Огайо. Журнал дикой природы Управление. 34 (4): 870-886. [16398] 71. Старый, Ричард. 1990. Камышевник обыкновенный (Chondrilla juncea L.) в Вашингтон: Идентификация, биология, экология и распространение. В: Рош, Бен Ф .; Рош, Синди Тэлботт, ред. Пересмотренные сорта сорняков: Труды симпозиума: Управление ареалом Тихоокеанского Северо-Запада 1989 г. краткий курс; 24-26 января 1989 г .; Спокан, Вашингтон.Пуллман, Вашингтон: Вашингтон Государственный университет, Департамент наук о природных ресурсах, Кооператив Добавочный: 71-76. [14839] 72. Radford, Albert E .; Ahles, Harry E .; Белл, К. Ричи. 1968. Наставник сосудистая флора Каролины. Чапел-Хилл, Северная Каролина: Университет Пресса Северной Каролины. 1183 с. [7606] 73. Raunkiaer, C. 1934. Жизненные формы растений и статистические растения. география. Оксфорд: Clarendon Press. 632 с.[2843] 74. Roberts, Teresa L .; Ванкат, Джон Л. 1991. Флористика хронопоследовательности. соответствует древней полевно-широколиственной последовательности лесов на юго-западе Огайо. II. Банки семян. Бюллетень Ботанического клуба Торри. 118 (4): 377-384. [17753] 75. Roland, A.E .; Смит, Э. С. 1969. Флора Новой Шотландии. Галифакс, NS: Музей Новой Шотландии. 746 с. [13158] 76. Schmidt, F. J. W. 1936. Зимняя пища острохвостого тетерева и перистый тетерев в Висконсине.Бюллетень Вильсона. Сентябрь: 186-203. [16729] 77. Schramm, J. R. 1966. Исследования колонизации растений на черных отходах от добыча антрацита в Пенсильвании. Сделки американского Философское общество. [Филадельфия, Пенсильвания]; 56 (1): 5-194. [24769] 78. Шифлет, Томас Н., изд. 1994. Типы пастбищных покровов США. Состояния. Денвер, Колорадо: Общество управления пастбищами. 152 с. [23362] 79. Skousen, J. G .; Колл, К.А.; Knight, R. W. 1990. Естественная растительность невостребованный бурый уголь на поверхности шахты в восточно-центральном Техасе. Юго-западный Натуралист. 35 (4): 434-440. [21195] 80. Стикни, Питер Ф. 1989. Одиночное происхождение видов, происходящих из северных Скалистые горные леса. Неопубликованный черновик находится в файле по адресу: Департамент США Сельское хозяйство, Лесная служба, Межгорная научно-исследовательская станция, Пожар Научная лаборатория, Миссула, штат Монтана; RWU 4403 файлов. 7 шт. [20090] 81.Стрейт, Ребекка А .; Джексон, Мэрион Т. 1986. Экологический анализ растительные сообщества природного заповедника Little Bluestem Prairie: предварительное сжигание по сравнению с последующим сжиганием. Слушания, Академия Индианы Наука. 95: 447-452. [22165] 82. Strausbaugh, P.D .; Core, Earl L. 1977. Флора Западной Вирджинии. 2-е изд. Моргантаун, Западная Вирджиния: Seneca Books, Inc. 1079 стр. [23213] 83. Sugihara, Neil G .; Рид, Лоис Дж .; Ленихан, Джеймс М. 1987. Растительность дубовые леса Лысых холмов, национальный парк Редвуд, Калифорния.Мадроно. 34 (3): 193-208. [3788] 84. Swales, Dorothy E. 1975. Необычная среда обитания для Drosera rotundifolia L., зимовку и вегетативное размножение. Канадский Полевой естествоиспытатель. 89 (2): 143-147. [22335] 85. Свон, Фредерик Р., младший, 1970. Постпожарная реакция четырех заводов. общины в южно-центральном штате Нью-Йорк. Экология. 51 (6): 1074-1082. [3446] 86. Свенсон, Джон Э. 1985. Сезонное использование среды обитания острохвостым тетеревом. Tympanuchus phasianellus, в прерии с разнотравьем в Монтане.Канадский Полевой естествоиспытатель. 99 (1): 40-46. [23501] 87. Тилман, Дэвид. 1988. Динамика и структура растительных сообществ. Монографии по популяционной биологии 26. Принстон, Нью-Джерси: Принстонский университет Нажмите. 360 с. [16944] 88. Служба охраны природных ресурсов Министерства сельского хозяйства США. 2018. База данных растений, [онлайн]. Служба охраны природных ресурсов Министерства сельского хозяйства США (производитель). Доступно: https://plants.usda.gov /. [34262] 89. Министерство внутренних дел США, Национальная биологическая служба. [нет данных]. НП Флора [База данных]. Дэвис, Калифорния: Министерство внутренних дел США, Национальное Биологическое обследование. [23119] 90. Ванкат, Джон Л .; Карсон, Уолтер П. 1991. Флористика хронопоследовательности. соответствующая успешности старых полево-лиственных лесов. на юго-западе Огайо. III. Пост-нарушенная растительность. Бюллетень ботанического сада Торри Клуб. 118 (4): 385-391.[17755] 91. Volland, Leonard A .; Делл, Джон Д. 1981. Влияние пожара на Тихий океан. Северо-западная лесная и пастбищная растительность. Портленд, Орегон: Департамент США Сельское хозяйство, Лесная служба, Тихоокеанский Северо-Западный регион, Управление пастбищами и авиация и управление пожарами. 23 п. [2434] 92. Восс, Эдвард Г. 1985. Флора Мичигана. Часть II. Двудольные (Saururaceae - Cornaceae). Бык. 59. Блумфилд-Хиллз, Мичиган: Крэнбрук. Институт Науки; Анн-Арбор, Мичиган: Гербарий Мичиганского университета.724 с. [11472] 93. Walters, R. S .; Ошмуди, Л. Р. 1989. Восстановление растительности на участок леса лиственных пород, оголенный рассолом. Ландшафт и градостроительство. 17: 127-133. [9819] 94. Weaver, T .; Lichthart, J .; Густафсон, Д. 1990. Экзотическое вторжение в лесная растительность, Северные Скалистые горы, США. В: Шмидт, Вайман C .; Макдональд, Кэти Дж., Составители. Труды - симпозиум по белой коры сосновые экосистемы: экология и управление высокогорным ресурсом; 29-31 марта 1989 г .; Bozeman, MT.Gen. Tech. Представитель INT-270. Огден, штат Юта: США Департамент сельского хозяйства, лесной службы, межгорных исследований Станция: 208-213. [11688] 95. Welsh, Stanley L .; Этвуд, Н. Дуэйн; Гудрич, Шерел; Хиггинс, Ларри С., ред. 1987. Флора Юты. Мемуары естествоиспытателя Большого бассейна № 9. Прово, UT: Университет Бригама Янга. 894 с. [2944] 96. Wilson, Scott D .; Тилман, Д. 1991. Интерактивные эффекты оплодотворения. и нарушение структуры сообщества и доступности ресурсов в старополевое растительное сообщество.Oecologia. 88: 61-71. [25193] 97. Винтерхолдер, Кит. 1990. Триггерный подход к инициированию. естественного возобновления растительных сообществ на промышленно поврежденных земли в Садбери, Онтарио. В: Хьюз, Х. Гленн; Бонниксен, Томас М., ред. Реставрация '89: новый вызов менеджменту: Труды, 1-е место ежегодное собрание Общества экологической реставрации; 1989 Январь 16-20; Окленд, Калифорния. Мэдисон, Висконсин: Дендрарий Висконсинского университета, Общество экологического восстановления: 215-226.[14697] 98. Уоффорд, Б. Юджин. 1989. Путеводитель по сосудистым растениям Голубого океана. Ридж. Афины, Джорджия: Издательство Университета Джорджии. 384 с. [12908] 99. Zimmerman, G.T .; Нойеншвандер, Л. Ф. 1984. Выпас скота. влияет на структуру сообщества, интенсивность пожаров и частоту пожаров в пределах типа среды обитания пихты Дугласа / девятикорной. Журнал диапазона Управление. 37 (2): 104-110. [10103] 100. Dittberner, Phillip L .; Олсон, Майкл Р.1983. Завод информации. база данных сети (PIN): Колорадо, Монтана, Северная Дакота, Юта и Вайоминг. FWS / OBS-83/86. Вашингтон, округ Колумбия: Министерство внутренних дел США, Служба рыбы и дикой природы. 786 с. [806] 101. Голли, Фрэнк Б. 1965. Структура и функции метлы на старых полях. сообщество. Экологические монографии. 35 (1): 113-137. [17419]
Домашняя страница FEIS
Щавель (Rumex acetosa L.): не только сорняк, но и перспективное овощное и лекарственное растение
Abbasi, A.М., М. Х. Шах, Т. Ли, Х. Фу, Х. Го и Р. Х. Лю. 2015. Этномедицинская ценность, фенольный состав и антиоксидантные свойства диких кулинарных овощей. Журнал этнофармакологии 162: 333-345.
CAS PubMed Google Scholar
Abbet, C., R. Mayor, D. Roguet, R. Spichiger, M. Hamburger & O. Potterest. 2014. Этноботаническое исследование дикорастущих альпийских пищевых растений в Нижнем и Центральном Вале (Швейцария). Журнал этнофармакологии 151: 624-634.
PubMed Google Scholar
Ахмад, С., Ф. Уллах, А. Садик, М. Айях, М. Имран, И. Али, А. Зеб, Ф. Уллах и М. Р. Шах. 2016. Химический состав, антиоксидантный и антихолинэстеразный потенциал эфирного масла Rumex hastatus D. Don, собранного на северо-западе Пакистана. BMC Дополнительная и альтернативная медицина 16: 29.
Эйнсворт, К., А. Рахман, Дж. Паркер и Г. Эдвардс.2005. Интерсексуальные соцветия Rumex acetosa демонстрируют, что определение пола уникально для каждого цветка. Новый фитолог 165: 711-720.
CAS PubMed Google Scholar
Али-Штайех, М.С., Р.М. Джамус, Дж. Х. Аль-Шафи, Вашингтон, Эльгераба, Ф. А. Херфан, К. Х. Карария, И. С. Хдаир, И. М. Сус, А. А. Муслех, Б. А. Иса, Х. М. Герзаллах, КБ Хлайф, С. М. Айаш, Г. М. Сва , М.А. Абузахра, М.М. Хадж-Али, Н.А. Сайфи, Х. К. Азем и Х. А. Насралла. 2008. Традиционные знания о дикорастущих съедобных растениях, используемых в Палестине (Северный Западный берег): сравнительное исследование. Журнал этнобиологии и этномедицины 4: 13.
PubMed PubMed Central Google Scholar
Alzoreky, N. S. & K. Nakahara. 2001. Антиоксидантная активность некоторых съедобных йеменских растений оценена с помощью анализа феррилмиоглобина / ABTS? +. Исследования пищевых продуктов и технологий 7: 141-144.
CAS Google Scholar
Андерсен, С. Т. 1990. Спектры пыльцы из двух подвешенных кувшинов раннего неолита в длинном кургане в Бьорнсхольме, Дания. Журнал датской археологии 9: 59-63.
Google Scholar
Bae, J-Y., Y. S. Lee, S. Y. Han, E. J. Jeong, M. K. Lee, J. Y. Kong, D. H. Lee, K. J. Cho, H-S. Ли и М.-Дж. Ан. 2012. Сравнение водного и этанолового экстрактов Rumex acetosa для защиты от язв желудка у мышей.Биомолекулы и терапия 20: 425–430.
Google Scholar
Beckert, S. & A Hensel. 2013. Протеиназо-ингибирующая активность экстракта Rumex acetosa L. против факторов вирулентности Porphyromonas gingivalis. Planta Medica 79: PJ6. https://doi.org/10.1055/s-0033-1352210
Белло О. М., П. С. Фасину, О. Э. Белло, А. Б. Огбесеяна, К. О. Адтунджи, А. О. Дада, О. С. Ибитое, С. Алоко и О. С.Огунтое. 2019. Дикий овощ Rumex acetosa Linn: этноботаника, фармакология и фитохимия — обзор. Южноафриканский журнал ботаники 125: 149-160.
CAS Google Scholar
Bicker, J., F. Petereit & A. Hensel. 2009. Проантоцианидины и производное флороглюцина из Rumex acetosa L. Fitoterapia 80: 483–495.
CAS PubMed Google Scholar
Блоцка-Вандас М., Э. Сливинска, А. Грабовска-Йоахимиак, К. Мусиал и А. Й. Иоахимиак. 2007. Развитие мужских гаметофитов и два разных класса ДНК пыльцевых зерен у растения Rumex acetosa L. с системой половых хромосом XX / XY1Y2 и соотношением полов смещенным по женскому признаку. Половое размножение растений 20: 171-180.
Google Scholar
Кардон, Д. 2007. Натуральные красители. Источники, традиции, технологии и наука. Публикации архетипа, Лондон.
Чекканти, К., М. Ланди, Л. Инкрокчи, А. Пардосси и Л. Гуиди. 2020. Пригодность Rumex acetosa L., выращенного на гидропонике, в качестве свежесрезанного продукта. Horticulturae 2020, 6: 4. https://doi.org/10.3390/horticulturae6010004
Couplan, F. 2009. Le Regal Vegetal. Plantes Sauvages Comestibles. Encyclopedie des Plantes Comestibles de l’Europe. Éditions Sang de la Terre, Париж.
Доган Ю., Недельчева А.,. Лучай, К.Драгулеску, Г. Стефков, А. Маглайлич, Дж. Фернер, Н. Папп, А. Хайдари, Б. Мустафа, З. Дайч-Стеванович и А. Пьерони. 2015. О важности листа: этноботаника сармы в Турции и на Балканах. Журнал этнобиологии и этномедицины 11: 26.
PubMed PubMed Central Google Scholar
Демирезер, Л. Э., А. Куруузун-Уз, И. Бергер, Х.-Ж. Schiewe & A. Zeeck. 2001. Состав антиоксидантных и цитотоксических агентов из природного источника: антрахиноны и дубильные вещества из корней Rumex Patientia .Фитохимия 58: 1213-1217.
CAS PubMed Google Scholar
Derksen, A., A. Hensel, W. Hafezi, F. Herrmann, T. J. Schmidt, C. Ehrhardt, S. Ludwig & J. Kühn. 2014. 3-O-галлоилированные процианидины из Rumex acetosa L. ингибируют прикрепление вируса гриппа А. PLOS ONE 9 (10): e110089. https://doi.org/10.1371/journal.pone.0110089
CAS Статья PubMed PubMed Central Google Scholar
Эрнст В.Х. О., Ф. Нолле, С. Кратц и Э. Шнуг. 2004. Аспекты экотоксикологии тяжелых металлов в районе Гарца — экскурсия с гидом. Lanbauforschung Völkenrode 54: 53-71.
CAS Google Scholar
Эткин, Н. Л. 1994. The Cull of the Wild, in: Eating on the wild side — The Pharmacologic, Ecologic and Social Implication on Using Noncultigens, (ed. N. L. Etkin) Arizona University Press, Tuscon.
Flora Europaea.1993. Том 1: от Lycopodiaceae до Platanaceae. Ред. Т. Г. Тутин, В. Х. Хейвуд, Н. А. Берджес, Д. Х. Валентин, С. М. Уолтерс, Д. А. Уэбб. Издательство Кембриджского университета, Кембридж, Великобритания.
Gescher, K., A. Hensel, W. Hafezi, A. Derksen & J. Kuhn. 2011. Олигомерные проантоцианидины из Rumex acetosa L. ингибируют прикрепление вируса простого герпеса типа 1. Противовирусные исследования 89: 9–18.
CAS PubMed Google Scholar
Glatthaar-Saalmüller, B., У. Рауххаус, С. Роде, Дж. Хауншильд и А. Заальмюллер. 2011. Противовирусная активность in vitro двух препаратов растительного лекарственного средства Синупрет® против вирусов, вызывающих респираторные инфекции. Фитомедицина 19: 1-7.
PubMed PubMed Central Google Scholar
Grant S., A. Houben, B. Vyskot, J. Siroky, W-H. Пан, Дж. Маккас и Х. Сэдлер. 1994. Генетика определения пола у цветковых растений. Генетика развития 15: 214-230.
Гупта, К. К., П. Гупта, П. Сингх, С. В. Сингх и С. Агарвал. 2006. Химия природных красителей. В : М. Дэниэл, С. Д. Бхаттачарья, А. Арья и В. М. Раоле (ред.). Природные красители: возможности и проблемы, стр. 7-34. Научные издательства, Джодхпур, Индия.
Google Scholar
Хуссейн, М., С. М. Раза и Х. Дж. Халид. 2015. Фармакологически механистическая основа традиционного использования Rumex acetosa при нарушениях моторики кишечника и рвоте.Бангладешский фармакологический журнал 10: 548-554.
Google Scholar
Ито, Х. 1986. Влияние противоопухолевых агентов из различных природных источников на систему метаболизма лекарств, фагоцитарную активность и систему комплемента у мышей, несущих саркому 180. Японский журнал фармакологии 40: 435-443.
CAS PubMed Google Scholar
Ярич, С., З. Попович, М.Мачуканович-Йоцич, Л. Джурджевич, М. Миятович, Б. Караджич, М. Митрович и П. Павлович. 2007. Этноботаническое исследование использования диких лекарственных трав с горы Копаоник (Центральная Сербия), Journal of Ethnopharmacology 111: 160-175.
PubMed Google Scholar
Jund, R., M. Mondigler, H. Steindl, H. Stammer, P. Stierna & C. Bachert. 2012. Клиническая эффективность сухого экстракта пяти лекарственных трав при остром вирусном риносинусите.Ринология 50: 417-426.
CAS PubMed Google Scholar
Хан, С. В. и С. Хатун. 2008. Этноботанические исследования некоторых полезных трав долин Харамош и Бугроте в Гилгите, северные районы Пакистана. Пакистанский журнал ботаники 40: 43-58.
Google Scholar
Kihara, H. & T. Ono. 1923. Цитологические исследования на Rumex L., I.Хромосомы Rumex acetosa L. Botanical Magazine Tokyo 37: 84-90.
Google Scholar
Корпелайнен, Х. 1991. Вариация соотношения полов и пространственная сегрегация полов в популяциях Rumex acetosa и R. acetosella (Polygonaceae). Систематика и эволюция растений 174: 183-195.
Google Scholar
Корпелайнен, Х.1992. Модели распределения ресурсов у мужских и женских растений Rumex acetosa и R. acetosella . Oecologia 89: 133-139.
PubMed Google Scholar
Корпелайнен, Х. 1993. Фенологические различия между популяциями и полами у многолетних видов Rumex acetosa . Acta Oecologica 14: 287-297.
Google Scholar
Корпелайнен, Х.1998. Лабильное выражение пола у растений. Биологические обзоры 73: 157-180.
Google Scholar
Корпелайнен, Х. 1999. Генетическая дифференциация скрещенных, широко распространенных и опыляемых ветром видов, Rumex acetosa (Polygonaceae). Botanisches Jahrbücher für Systematik, Pflanzengeschichte und Pflanzengeographie 121: 269-277.
Google Scholar
Корпелайнен, Х.2002. Генетический метод определения пола дополняет исследования соотношений полов в Rumex acetosa . Молекулярная экология 11: 2151-2156.
CAS PubMed Google Scholar
Кота С., П. Думпала, Р. К. Ананта, М. К. Верма и С. Кандепу. 2017. Оценка терапевтического потенциала наночастиц серебра / хлорида серебра, синтезированных с водным экстрактом листьев Rumex acetosa . Исследования природы, научный отчет 7: 11566.https://doi.org/10.1038/s41598-017-11853-2
CAS Статья Google Scholar
Kotzamani, G. & A. Livarda. 2018. Запутывание людей и растений на заре сельскохозяйственной практики в Греции. Анализ археоботанических останков эпох мезолита и раннего неолита. Четвертичный международный 496: 80-101.
Google Scholar
Кубяк-Мартенс, Л.1999. Компонент растительной пищи в рационе поселения позднего мезолита (Эртеболле) в Тайбринд Виг, Дания. История растительности и археоботаника 8: 117-127.
Google Scholar
Кучекова З., Дж. Млчек, П. Гумполичек, О. Роп, П. Валасек и П. Саха. 2011. Фенольные соединения из Allium schoenoprasum , Tragopogon pratensis и Rumex acetosa и их антипролиферативные эффекты. Молекулы 16: 9207–9217.
CAS PubMed PubMed Central Google Scholar
Ladeji O. & Z.S.C. Окое. 1993. Химический анализ листа щавеля ( Rumex acetosa ). Пищевая химия 48: 205-206.
CAS Google Scholar
Ли Н. Дж., Дж. Х. Чой, Б.С. Ку, S-Y. Рю, YH. Хан, С.Дж. Ли и Д-У Ли. 2005. Антимутагенность и цитотоксичность компонентов из надземных частей Rumex acetosa .Биологический и фармацевтический бюллетень 28: 2158-2161.
CAS PubMed Google Scholar
Лидделл, Х.Г. и Скотт, Р. 1940. Греческий английский лексикон. 9 -е издание . Издательство Оксфордского университета. Ллойд, Д. 1982. Селекция комбинированных и отдельных полов семенных растений. Американский натуралист 120: 571-585.
Lloyd, D.G. 1982. Селекция комбинированных и отдельных полов семенных растений.Американский натуралист 120: 571-585.
Łuczaj, Ł. И В. М. Шиманский. 2007. Дикие сосудистые растения, собранные для употребления в польской деревне: обзор. Журнал этнобиологии и этномедицины 3: 17. 10.1186 / 1746-4269-3-17
Мабберли, Д. Дж. 2008. Книга растений Мабберли. 3 -е издание . Издательство Кембриджского университета, Кембридж
Google Scholar
Макаров, В., С. Макарова, А.Дж. Любовь, О. Синицына, А.О. Дудник, И.В. Яминский, М.Е. Тальянский, Н.О. Калинина. 2014. Биосинтез стабильных наночастиц оксида железа в водных экстрактах растений Hordeum vulgare и Rumex acetosa . Langmuir 30: 5982-5988.
CAS PubMed Google Scholar
Мантл, Д., Ф. Эддеб и А. Т. Пикеринг. 2000. Сравнение относительной антиоксидантной активности британских лекарственных растений in vitro.Журнал этнофармакологии 72: 47-51.
CAS PubMed Google Scholar
Мансано С., З. Мегиас, К. Мартинес, А. Гарсия, Э. Агуадо, Т. Чиле, Д. Лопес-Алонсо, Ф. Гарсия-Марото, Э. Кейновски, Я. Широки, З. Кубат, Т. Кралова, Б. Выскот и М. Джамилена. 2017. Сверхэкспрессия аэролизиноподобного белка, специфичного для цветка, из двудомного растения Rumex acetosa изменяет развитие цветов и вызывает мужскую стерильность у трансгенного табака.Журнал растений 89: 58-72.
CAS PubMed Google Scholar
Мариотти Б., С. Манзано, Э. Кейновски, Б. Выскот и М. Джамилена. 2009. Накопление Y-специфических сателлитных ДНК в ходе эволюции половых хромосом Rumex acetosa . Молекулярная генетика и геномика 281: 249. https://doi.org/10.1007/s00438-008-0405-7
CAS Статья PubMed Google Scholar
Мацухиса С.И А. Усимару. 2019. Существует ли половой диморфизм в фенологических признаках цветения у анемофильных раздельнополых видов? Тест с Rumex acetosa . Американский журнал ботаники 106: 1356-1364.
CAS PubMed Google Scholar
Mattalia, G., C. L. Quave & A. Pieroni. 2013. Традиционное использование дикорастущих продуктов питания и лекарственных растений в общинах бригасков, кие и провансальцев в западных итальянских Альпах. Генетические ресурсы и эволюция сельскохозяйственных культур 60: 587–603.
Google Scholar
Маклейн, Р. Дж., П. Т. Херли, М. Р. Эмери и М. Р. По. 2014. Сбор «дикой» пищи в городе: переосмысление роли добычи в планировании и управлении городской экосистемой. Международный журнал правосудия и устойчивости 19: 220-240.
Google Scholar
Mekonnen, T., K. Urga & E. Engidawork. 2010. Оценка мочегонной и анальгетической активности корневищ Rumex abyssinicus Jacq.у мышей. Журнал этнофармакологии 127: 433-439.
PubMed Google Scholar
Навахас-Перес, Р., Р. де ла Эрран, Г. Лопес Гонсалес, М. Джамилена, Р. Лосано, К. Р. Рейон, М. Р. Рейон и М. А. Гарридо-Рамос. 2005. Эволюция репродуктивных систем и механизмов определения пола в пределах Rumex (Polygonaceae), выведенная на основе данных ядерных и хлоропластидных последовательностей. Молекулярная биология и эволюция 2: 1929–1939.
Google Scholar
Недельчева А. 2013. Этноботаническое исследование дикорастущих съедобных растений Болгарии. Евразийский журнал бионаук 7: 7-94.
Google Scholar
Noonan, S.C. & G.P. Savage. 1999. Содержание оксалатов в пищевых продуктах и его влияние на человека. Азиатско-Тихоокеанский журнал клинического питания 8: 64-74.
CAS PubMed Google Scholar
Оно, Т.1935. Chromosomen und Sexualität von Rumex acetosa . Научные отчеты Императорского университета Тохоку IV 10: 41-210.
Google Scholar
Пардо де Сантаяна, М., Х. Тардио, Э. Бланко, А. М. Карвалью, Х. Дж. Ластра, Э. Сан, М. Моралес и Р. Моралес. 2007. Традиционные знания о диких съедобных растениях, используемых на северо-западе Пиренейского полуострова (Спин и Португалия): сравнительное исследование. Журнал этнобиологии и этномедицины 3: 1-11.
Google Scholar
Паркер, Дж. С. 1990. Половая хромосома и половая дифференциация у цветковых растений. Хромосомы сегодня 10: 187-198.
CAS Google Scholar
Паркер, Дж. С., М. С. Кларк. 1991. Дозировка систем половых хромосом у растений. Наука о растениях 80: 79–92.
Google Scholar
Паркер П.2018. Маленькая тетрадь на латыни для садоводов. Литтл, Браун, Лондон.
Google Scholar
Перейра, К., Л. Баррос, А. М. Карвалью и К. Ф. Р. Феррейра. 2013. Использование УФЖХ-КПК для анализа органических кислот в тридцати пяти видах пищевых и лекарственных растений. Методы анализа пищевых продуктов 6: 1337-1344.
Google Scholar
Пьерони, А., К. Чианфальоне, А. Недельчева, А.Хайдари, Б. Мустафа и К. Quave. 2014. Устойчивость на границе: традиционные ботанические знания македонцев и албанцев, проживающих в Голлобордо, Восточная Албания. Журнал этнобиологии и этномедицины 10: 31. https://doi.org/10.1186/1746-4269-10-31
Пьерони А. и М. Э. Джусти. 2009. Альпийская этноботаника в Италии: традиционные знания о гастрономических и лекарственных растениях у окситанцев верхней долины Вараита, Пьемонт. Журнал этнобиологии и этномедицины 5:32.https://doi.org/10.1186/1746-4269-5-32
Камар, Х. М. У. Д., Р. Кайюм, У. Салма, С. Хан, Т. Хан и А. Дж. Шах. 2018. Сосудистые механизмы, лежащие в основе гипотензивного действия Rumex acetosa . Фармацевтическая биология 56: 225-234.
PubMed PubMed Central Google Scholar
Куреши, Р. А., М. А. Гуфран, К. Н. Султана, М. Ашраф и А. Г. Хан. 2007. Этномедицинские исследования лекарственных растений Гилгитского района и прилегающих территорий.Приложения исследований этноботаники 5: 115–122.
Google Scholar
Салех Н., М. Н. Эль-Хадиди и Р. Ф. М. Арафа. 1993. Флавоноиды и антрахиноны некоторых видов египетских Rumex (Polygonaceae). Биохимическая систематика и экология 21: 301-303.
CAS Google Scholar
Samancioglu, A., I.G.Sat, E. Yildirim, S. Ercisli, T. Jurikova & J.Млчек. 2016. Оценка общего содержания фенолов и витамина С и антирадикальной активности в традиционно потребляемых диких съедобных овощах из Турции. Индийский журнал традиционных знаний 15: 208-213.
Google Scholar
Schmuch, J. M., S. Beckert & A. Hensel. 2013. Проантоцианидины из Rumex acetosa L. увеличивают in vitro скорость фагоцитоза Porphyromonas gingivalis в макрофагах мыши и обеспечивают цитопротекторный и противовоспалительный потенциал для современной терапии пародонтита.Planta Medica 79 — SL15.
Шибата, Ф., М. Хидзуме и Ю. Куроки. 1999. Хромосомная окраска Y-хромосом и выделение специфической повторяющейся последовательности Y-хромосомы в двудомном растении Rumex acetosa . Хромосома 108: 266–270.
CAS PubMed Google Scholar
Шибата, Ф., М. Хидзуме и Ю. Куроки. 2000. Дифференциация и полиморфная природа Y-хромосом, выявленная с помощью повторяющихся последовательностей у двудомного растения, Rumex acetosa .Исследование хромосом 8: 229–236.
CAS PubMed Google Scholar
Сыуканд, Р. и А. Пьерони. 2019. Устойчивость в горах: биокультурные убежища дикой пищи в Большом Кавказском хребте, Азербайджан. Биоразнообразие и сохранение 28: 3529–3545.
Google Scholar
Sun, Y. Y., X. H. Su & J. Y. Jin. 2015. Rumex acetosa L.индуцирует вазорелаксацию в аорте крысы посредством активации передачи сигналов PI3-киназы / Akt- И Ca2 + -eNOS-NO в эндотелиальных клетках. Журнал физиологии и фармакологии 66: 907-915.
CAS PubMed Google Scholar
Tamokou de Dieu J., JR Chouna, E. Fischer-Fodor, G. Cherches, O. Barbos, D. Benedec, M. Duma, APN Efouet, HK Wabo, JR Kuiate, A. Mot & R. Силаги-Румитреску. 2013. Противоопухолевое и противомикробное действие некоторых богатых антиоксидантами камерунских лекарственных растений.PLoS ONE 8 (2): e55880. https://doi.org/10.1371/journal.pone.0055880
Танака, Т. 1976. Циклопедия съедобных растений мира Танаки. Издательство Кэйгаку, Токио.
Google Scholar
Тейлор Р. С. Л., Ф. Эдель, Н. П. Манандхар и Г. Х. Н. Тауэрс. 1996. Антимикробная активность лекарственных растений южного Непала. Journal of Ethnopharmacology 50: 97-102
Tolrà, R.P., C.Пошенридер, Б. Луппи и Дж. Барсело. 2005. Вызванные алюминием изменения профилей как органических кислот, так и фенольных веществ лежат в основе толерантности к Al в Rumex acetosa L. Environment and Experimental Botany 54: 231-238.
Google Scholar
Туазон-Нартеа, Дж., Дж. Сэвидж. 2013. Исследование уровней оксалатов в частях растений щавеля и продуктах на его основе. Науки о пищевых продуктах и питании 4: 838-843.
Google Scholar
Ванханен, С.И П. Песонен. 2016. Сбор дикорастущих растений в Финляндии каменного века. Четвертичный международный 404, А: 43-55.
Google Scholar
Vasas, A. ,, O. Orbán-Gyupai & J. Hohmann. 2015. Род Rumex . Обзор традиционных применений, фитохимии и фармакологии. Журнал этнофармакологии 175: 198-228.
CAS PubMed Google Scholar
Ван, К.Р., Ю. С. Цуй, Х. Лю, Ю. Т. Донг и П. Кристи. 2003. Загрязнение почвы и поглощение тяжелых металлов растениями на загрязненных участках в Китае. Журнал экологической науки и здоровья A, 38: 823-838.
Google Scholar
Уивер, W. W. 2000. 100 овощей и их происхождение. Алгонкинские книги Чапел-Хилл, Чапел-Хилл, Северная Каролина, США.
Wegiera, M., H. D. Smolarz, D. Wianowska & A. L. Dawidowicz. 2007 г.Производные антрацена у некоторых видов рода Rumex L. Acta Societatis Botanicorum Poloniae 76: 103-108.
CAS Google Scholar
Wegiera, M., H. D. Smolarz и A. Bogucka-Kocka. 2012. Rumex L. виды индуцируют апоптоз в клеточных линиях 1301, EOL-1 И H-9. Acta Poloniae Pharmaceutica — Исследования лекарственных средств, 69: 487-499.
Google Scholar
WFO.2020. Мировая флора онлайн. Опубликовано в Интернете; http://www.worldfloraonline.org. По состоянию на 1 июля 2020 г.
Xiong, J. 1998. Использование гибрида Rumex acetosa L. в производстве продукта, содержащего пшеничную муку. Патент WO199
84A1.
Ye, M., B. Liao, J. T. Li, A. Mengoni, M. Lu, W. C. Luo, W. S. Shu. 2012. Контрастные паттерны генетической дивергенции у двух симпатрических псевдометаллофитов: Rumex acetosa L. и Commelina communis L.BMC Evolutionary Biology 12:84.
Чжоу, X., Г. X. Ван и Ф. Ян. 2012. Удаление азота из эвтрофных речных вод с использованием Rumex acetosa , выращиваемого в экологических плавучих руслах. Бюллетень по окружающей среде Fresenius 21: 1920-1928.
CAS Google Scholar
Овца-щавель
Овечий щавель
Румекс Ацетоселла
Кислая трава, щавель полевой, щавель красный верховой, щавель садовый, соус зелень, kliener ampfer, oseille
Многолетнее травянистое растение семейства гречиховых ( Polygonaceae )
вырастает до высоты от 4 до 12 дюймов.Стебли прямостоячие, на верхушке ветвистые, тонкие, красноватого цвета. Зеленые листья в форме стрелы простые, 1-3 дюйма в длину и гладкие с парой горизонтальных лопастей у основания. Нижние листья лопатообразные, без лопастей. Цветки от зеленого до красного до ржаво-коричневого и собираются в группы около верхушки растения. Мужские и женские цветки обычно находятся на разных растениях, мужские цветки от желтого до красного, а женские — зеленоватые. Цветут с мая по октябрь. Семена красноватые или золотисто-коричневые с ржаво-коричневой оболочкой, которая прилипает к семенам.Семена могут оставаться всхожими в почве от 10 до 20 лет. Корневая система
состоит из мелких волокнистых корней и обширных горизонтальных корней, которые могут достигать глубины 5 футов. Щавель овец размножается семенами и ползучими корнями, дающими новые побеги.
Овечий щавель содержит витамины A, B, C, D, E, K, минералы кальций, хлор, железо и магний. Кроме того, овечий щавель содержит небольшое количество натрия, серы, кремния и следы меди,
йод, марганец и цинк.Сушеные надземные части щавеля овец содержат:
рутин (0,53%), флавоновые гликозиды (т.е. гиперозид или
кверцитин-3d-галактозид) 0,05% и гиперин (12 мг / 100 г). Общий витамин С
листьев варьируется от 750 до 1200 мг / 100 г в пересчете на сухой вес. Ясень
(8,1%) содержит в форме оксида 20,0% кальция; 13,9% фосфора;
13,4% магния; 28,3% калия и 11,5% кремния, а также железо, сера, медь, йод, марганец и цинк. Листья и стебли содержат полезные каротиноиды, хлорофилл, органические кислоты (т.е., яблочный,
щавелевая, дубильная, винная и лимонная) и фитоэстрогены. Растение также содержит поддерживающие здоровье антрахиноны, включая эмодин, алоэ-эмодин, хризофанол, реин и физсион.
- Стимулирует выведение мочи (мочегонное средство)
- Стимулирует транспирацию (потогонное)
- Тоник для почек и мочевыводящих путей
- Освежающий утоляющий жажду
- Уменьшение выработки желчи
- Бодрит сердце во время лихорадки
- Противодействует гниению
- Укрепляет иммунную систему
- Укрепляет клеточные стенки
- Помогает очищать кровеносные сосуды
- Стимулирует рост новых тканей
- Повышает содержание кислорода в клетках
Исторически или традиционно известно, что поддерживает функции и физиологию организма как:
- Вяжущее
- Противовоспалительное
- Антиоксидант
- Регенерация клеток
- Очищение
- Детоксикация
- Заболевания десен (противоскорбутиковые)
- Мочегонное слабительное
- Увеличение лимфатических узлов
- Сосудистые заболевания
- Лихорадка (жаропонижающее)
- Золотуха
- Сердечно-сосудистая деятельность
- Мягкий антисептик
- Мочегонное
- Питательный
Прием больших доз овечьего щавеля может вызвать желудочный
нарушение, тошнота и диарея из-за слабительного антрахинонового типа
соединения.Большие дозы сырых трав могут даже вызвать отравление из-за:
высокое содержание щавелевой кислоты и танинов.
Щавель овечий и другие растения семейства Polygonaceae содержат
оксалаты в свежем виде и оксалаты в приготовленных листьях (аналогичные
к шпинату или ревеню) и может быть противопоказан при заболеваниях почек
камни или слабая функция почек.
Щавель овечий в основном применяется в виде отвара (экстракция
эссенция или активный ингредиент из вещества путем кипячения).Но его также можно употреблять в сыром виде, например, в салате. Семена имеют
детоксицирующий эффект. Из-за присутствия щавелевой кислоты не следует превышать рекомендуемое количество.
Исторические архивы народной медицины в Европе и США неоднократно упоминают овечий щавель. Еще в 1740 г. трава для пациентов была разрешена законом.
Медсестра Рене Кайсс, известная пользователь группы травяных смесей, которую она назвала Essiac, считала овечий щавель ключевой травой в протоколе разрушения нежелательных тканей и лишних веществ в организме.Овечий щавель также активировал детоксикацию и очистку клеток, но работал лучше в синергии с другими травами в смеси.
Щавель овечий содержит много хлорофилла, зеленого пигмента крови растений, и способствует переносу большого количества кислорода в клетки. Это может быть замечательно, помогая поддерживать целостность клеток в ситуационных открытых повреждениях клеток (вызванных, например, радиацией от использования рентгеновских лучей). Также были продемонстрированы мощные антиоксидантные эффекты каротиноидов в овечьем щавеле.
Щавель овечий обеспечивает кислородом ткани на глубоком клеточном уровне. Это также обеспечивает сильную структурную иммунную поддержку. Овечий щавель имеет давнюю историю традиционного использования в качестве вяжущего, мочегонного и мягкого слабительного средства. Овечий щавель содержит такие компоненты, как бета-каротин, винный камень кислота, оксалаты (щавелевая кислота), антрахиноны (хризофанол, эмодин и rhein), гликозиды, такие как гиперозид, кверцитин-3d-галактозид. В растение родом из Евразии, но было завезено в большинство остальных Северной Америки считается обычным сорняком на полях, луга и леса.
По крайней мере десять коренных племен Канады и Соединенных Штатов использовали это растение, также известное как кислая трава или кислый сорняк, в пищу и медицина. Овечий щавель является популярным ингредиентом многих народных средств, а чай традиционно использовался как мочегонное средство и для снятия лихорадки. воспаления и заболевания десен. Интересно, что хоть это и не бобовые, овечий щавель содержат значительный уровень фитоэстрогенов с заметная активность связывания рецептора эстрогена, аналогичная изофлавону фитоэстрогены, общие для красного клевера, солодки и сои, всех бобовых культур, известных своими сильными восстанавливающими свойствами.Трава также содержит несколько антрахинонов, которые являются эффективными антиоксидантами и поглотителями свободных радикалов. Хотя исследования овечьего щавеля ограничены, близкородственные виды содержат мощное антибактериальное соединение, называемое румицином.
Высокое содержание танинов в чае также оказывает вяжущее действие, который полезен при таких проблемах, как диарея и чрезмерное менструальное кровотечение.
Высокое содержание танинов в чае также может обладают вяжущим действием, что полезно при таких проблемах, как диарея и обильное менструальное кровотечение.При низких дозах большинство видов Rumex полезен при диарее; однако в более высоких дозах они стимулируют перистальтика из-за наличия антрахинонов, которые непосредственно влияют на нервно-мышечной ткани, увеличивая слизистую продукцию толстой кишки клетки слизистой оболочки и способствуют секреции воды в кишечник просвет, тем самым оказывая очищающий эффект.
Rumex acetosella — Bugwoodwiki
Автор этой аннотации: Марк Фрей, Presidio Trust
Таксономия
Королевство:
Plantae
Тип:
Magnoliophyta
Класс:
Magnoliopsida
Род: Rumex Виды: R.acetosella Подвиды: R. acetosella Научное название Rumex acetosella соррель
L.
- Внешний вид
- Rumex acetosella — это многолетнее растение, которое может достигать 0,5 м в высоту.
- Листва
- Листья очередные, черешковые, голые, цельнокрайние, трехлопастные, до 1.2 дюйма (3 см) в длину и расположены в базальной розетке.
- Цветы
- Цветение происходит с марта по ноябрь, когда желтовато-зеленые (мужские) или красноватые (женские) цветки развиваются в гроздьях разветвленных соцветий на вершине стебля. Цветки, как правило, поникшие, имеют 3 лепестка и 3 чашелистика, каждый менее 0,04-0,06 дюйма (1-1,5 мм) в длину.
- Фрукты
- Плоды — красные семянки.
- Экологическая угроза
- Rumex acetosella произрастает в Европе и встречается по обочинам дорог и других нарушенных территориях.
Rumex acetosella L.
- ITIS (2007) перечисляет три внутривидовых таксона, но отмечает, что они не принимаются [1] .
- Rumex acetosella ssp. angiocarpus (Murb.) Murb.
- Rumex acetosella var. pyrenaeus (Pourret) Тимбал-Лаграв
- Rumex acetosella var. tenuifolius Wallr.
СИНОНИМЫ
Следующие синонимы перечислены в IPANE (2007) [2] :
- Acetosella acetosella (L.) Маленький
- Acetosella tenuifolia (Wallr.) A. Löve
- Acetosella vulgaris (Koch) Fourr.
- Rumex acetosella ssp. angiocarpus (Murb.) Murb.
- Rumex acetosella var. pyrenaeus (Pourret) Тимбал-Лаграв
- Rumex acetosella var. tenuifolius Wallr.
- Rumex angiocarpus Murb.
- Rumex tenuifolius (Wallr.) A. Löve
ОБЩЕЕ НАЗВАНИЕ
Распространенные названия Rumex acetosella включают овечий щавель, полевой щавель и красный щавель.
ОПИСАНИЕ И ДИАГНОСТИЧЕСКИЕ ХАРАКТЕРИСТИКИ
Rumex acetosella — двудомное травянистое многолетнее растение с ползучими корневищами. Его высота 10-40 см (4-16 дюймов) [3] . Корни могут достигать глубины 1,5 м (5 футов) [2] . Стебли узкие, красноватые.
Листья 2-10 см (0,8-4 дюйма)) в длину и 1-2 см (от 0,4 до 0,8 дюйма) в ширину [4] . Большинство листьев расположены в прикорневой розетке, но часто бывают стеблевые листья. Прикорневые листья очередные и лопастные, с конечной ланцетной частью (2-12 мм (0,08-0,5 дюйма) в длину), а боковые лопасти намного меньше и треугольной формы (стреловидные или хастатные основания листа). Прикорневые листья примерно 2,5-7,6 см (1-3 дюйма) в длину, черешковые и голые. Стеблевые листья очередные, обычно без лопастей, без черешков или с небольшими черешками [5] .Все листья имеют тонкую перепончатую оболочку (раструб), окружающую стебель в месте прикрепления черешка. Стреловидные листья и отчетливый красноватый оттенок являются полезными отличительными характеристиками.
Корни желтоватые, от очень тонких (<0,5 мм) до больших (> 3 мм) в диаметре (личное наблюдение).
Цветущие стебли могут быть в два раза короче общей длины растения. Цветки у мужских и женских растений морфологически различаются. Мужские цветки желтые, женские — красноватые [6] ).Цветки кивают на коротких сочлененных цветоножках. Мужские цветки имеют обратнояйцевидные внутренние листочки околоцветника размером 1,5-2 мм (0,06-0,08 дюйма). У женских цветков листочки околоцветника широко-яйцевидные [2] .
Блестящие золотисто-коричневые плоды — это семянки, трехугольные, размером примерно 1,5 мм (0,06 дюйма). В Новой Англии плоды присутствуют на растении с июня по октябрь [2] .
Семядоли безволосые, продолговатые, до 10 мм (0,4 дюйма) в длину.Семядоли и молодые листья не имеют гладкой поверхности. Молодые листья продолговатые; базальные стреловидные листья развиваются позже [5] .
ПОДОБНЫЕ ВИДЫ
В роду [7] насчитывается 200 видов. Rumex acetosa L. (щавель садовый) аналогичен, но более крупный, чем R. acetosella , [2] (Delmas 1993). В США R. acetosa зарегистрирован из Коннектикута, Массачусетса, Мэна, Мичигана, Миннесоты, Монтаны, Нью-Гэмпшира, Нью-Джерси, Нью-Йорка, Орегона, Пенсильвании, Вермонта, Вашингтона, Висконсина и Вайоминга [8] .
ОБЩАЯ ИНФОРМАЦИЯ
Удаление биомассы, особенно удаление корней, является ключом к успеху. Сеянцы, фрагменты корней, оставленные под землей, и любые части растений, оставленные над землей, могут быть источником новых растений. Его корни могут достигать глубины 1,5 м (5 футов), что затрудняет контроль и управление с помощью ручных и / или механических методов. Этот вид чувствителен к глифосату и гербицидам, содержащим дикамбу.
ОБИТАЙ И АССОРТИМЕНТ
Rumex acetosella произрастает в большинстве стран Европы, России, Ближнего Востока и Северной Африки [2] . R. acetosella натурализован в Новой Зеландии, Гренландии, западной части Южной Америки, южной оконечности Африки, Исландии (GBIF 2007) и США [8] . В США это происходит во всех 50 штатах [8] . APWG (2007) перечисляет его как инвазивный в Аризоне, Коннектикуте, Гавайях, Нью-Йорке, Орегоне, Теннесси, Вирджинии, Вашингтоне, Висконсине и Западной Вирджинии. Rumex acetosella был назван ядовитым сорняком в Айове [2] и запрещен к ввозу и продаже в Коннектикуте (ЗАВОДЫ USDA).Этот вид встречается в самых разных средах обитания, но особенно в полях и нарушенных местах. Обычен в поймах рек [9] , дубовых лесах и желтых сосновых лесах [6] .
Открытые площадки
Rumex acetosella часто встречается на пастбищах или в других открытых местообитаниях. В западном Вашингтоне встречается в районах, где преобладает Salix scouleriana; он также растет с Fragaria virginiana , Holcus lanatus , Trifolium repens , Rumex crispus и Juncus effusus [9] (Fonda 1974).В Орегоне R. acetosella обычно ассоциируется с Bromus tectorum [9] . В Коннектикуте он встречается на пастбищах Schizachyrium scoparium , выращиваемых после сельскохозяйственных культур, с Agrostis alba и Carex pensylvanica [9] (Niering and Dreyer 1989). В Нью-Джерси он растет с Solidago canadensis [9] (Carson and Picket 1990). Произрастает в лысых травянистых сообществах южных Аппалачей, где преобладают виды Danthonia compressa и Rubus canadensis , Vaccinium pallidum и Viola spp. [9] (Линдси и др., 1979, Марк, 1958). В Индиане R. acetosella встречается с Schizachyrium scoparium , Celtis occidentalis и Equisetum laevigatum [9] (Strait and Jackson 1986). В Монтане и Вайоминге он растет в альпийской тундре [9] (Weaver et al. 1990). В Южной Каролине растет R. acetosella в нарушенном состоянии. Andropogon virginicus [9] (Golley 1965). В Калифорнии было отмечено произрастание в пресноводных болотах с Festuca arundinacea , Carex spp., Typha angustifolia [9] (Fiedler and Leidy, 1987). В Калифорнии R. acetosella обычен на однолетних пастбищах, горных лугах и многолетних кустарниковых сообществах. Ассоциированные компании включают Bromus Rigidus , B. hordeaceus , Aira caryophyllea , Poa pratense , P. nevadensis , Carduus pycnocephalus , Avena fattifua 9023 [9] Boyd et al.1993, Дэвис и Шерман 1992, Фидлер и Лейди 1987, Макбрайд и др. 1991). На национальном побережье Пойнт-Рейес, Калифорния, R. acetosella встречается в сообществе прибрежных пастбищ с прибрежным каменным кресс-салатом Arabis blepharophylla , Toxicodendron diversilobum , Berberis pinnata и Chorizanthe valida и Fellers 1986, Davis and Sherman 1992).
Лесные массивы
Rumex acetosella также можно найти в лесных районах, включая следующие.В Пенсильвании R. acetosella встречается в сообществах Pinus strobus — Danthonia spicata с Solidago canadensis , Epilobium angustifolium , Lysimachia quadrifolia , Claytonia virginica и Viola spp. [9] (Ошмуди и Уолтерс 1988, Колб и др. 1990, Уолтерс и Очмуди 1989). В Альберте, R. acetosella является представителем 80-летнего растения Picea glauca — Pinus Banksiana — Pleurozium spp.community [9] (Fyles 1989). В Айдахо R. acetosella встречается с Abies grandis , Asarum caudatum , Pachistima myrsinites и Physocarpus malvaceus [9] (Грин, Дженсен и др., 1985, Leege и Годб. , Zimmerman and Neunschwander 1984). В Калифорнии R. acetosella встречается в Sequoia sempervirens , Pseudotsuga menziesii , Quercus garryana местообитаниях [9] (Gardner 1958, Krueger and Donart 1974, Sugihara et al.1987). В предгорьях Сьерра-Невады и Прибрежных хребтов, Калифорния, R. acetosella растет под Quercus douglasii [9] (Голландия, 1980).
ВОЗДЕЙСТВИЯ И УГРОЗЫ, ПРЕДСТАВЛЕННЫЕ Rumex acetosella
Он может образовывать колонии диаметром от 10 до 30 футов, за исключением других растений [6] .
БИОЛОГИЯ И ЭКОЛОГИЯ
Свет и температура
R. acetosella растет в различных условиях, но, по-видимому, предпочитает открытые участки (Parker 1995).Он легко переносит морозы зимой в северных штатах.
Состояние почвы (pH, влажность, текстура, питательные вещества)
Rumex acetosella предпочитает районы с очень бедной кислой почвой с низким содержанием азота. Плохо растет на известковых почвах [2] . В одном исследовании пиковая плотность наблюдалась при низком содержании азота (Tilman 1988). В Западной Вирджинии R. acetosella хорошо растет во всех регионах, кроме известняковых [9] (Strausbaugh and Core 1977).
Устойчивость к наводнениям и / или засухе
Неизвестно
Репродукция
Размножаются как семенами, так и вегетативно. Особи часто образуют огромные популяции с одним или несколькими генами.
Даты цветения:
| Государственный | месяцев | ссылку |
|---|---|---|
| Грузия | март-июнь | (Wofford 1989, цитируется по Esser 1995) |
| Теннесси | март-июнь | (Wofford 1989, цитируется по Esser 1995) |
| Вирджиния | март-июнь | (Wofford 1989, цитируется по Esser 1995) |
| Северная Каролина | март-июль | (Radford et al.1968 и Wofford 1989, оба в Esser 1995) |
| Южная Каролина | март-июль | (Radford et al.1968 и Wofford 1989, оба в Esser 1995) |
| Калифорния | март-август | (Munz 1973) |
| Канзас | апрель-июль | (Bare 1979, цитируется по Esser 1995) |
| Великие равнины | апрель-август | (Ассоциация флоры Великих равнин 1986, цитируется по Esser 1995) |
| Северная Дакота | май-июнь | (Диттбернер и др.1983, цитируется по Esser 1995). |
| Монтана | май-август | (Диттбернер и др. 1983, цитируется по Esser 1995) |
| Айдахо | май-сентябрь | (Фитцсиммонс и Беррилл, 1993) |
| Орегон | май-сентябрь | (Фитцсиммонс и Беррилл, 1993) |
| Вашингтон | май-сентябрь | (Фитцсиммонс и Беррилл, 1993) |
| Западная Вирджиния | май-сентябрь | (Strausbaugh and Core 1977, цитируется по Esser 1995) |
р.acetosella быстро распространяется, давая новые растения из придаточных стеблевых почек на корнях. Эти почки обычно находятся в верхних 20 см (8 дюймов) слоя почвы [9] (Kiltz 1930).
Щавель овечий вторгается в нарушенные участки и может переходить на нетронутые участки. Он быстро колонизируется семенами и может сохраняться от 15 до 20 лет (Escarre et al. 1994). Конкуренция с другими видами на хороших почвах может снизить его численность (Fitzsimmons and Burrill 1993).
В 2006 году в Президио Сан-Франциско было проведено тепличное исследование для оценки выживаемости 5 дюймовых.(12,7 см) R. acetosella Фрагменты корней разной толщины при заглублении на разную глубину. Со временем более глубоко закопанные корни дали листья. Через месяц все корни, оставшиеся на поверхности, за исключением самых толстых, дали листья. Через два месяца все корни, заглубленные на два дюйма, также дали листья. Через три месяца самые толстые корни, заглубленные на глубину четырех дюймов, также дали листья. Через три месяца растения не прорастали. Нет корней, заглубленных на глубине 6 дюймов и более, при появлении листьев (Frey et al.2008 г.).
Опыление
Опыление ветром.
Производство семян
Неизвестно
Распределение семян, Банк семян
Семена разносятся ветром и насекомыми [2] . Блестящие трехугольные семянки красного дерева и длиной 1,6 мм (0,063 дюйма) или менее [6] . Овечий щавель имеет устойчивый семенной фонд и может присутствовать в виде семян даже при отсутствии растущих растений. Эти семена прорастают после нарушения [9] (Del Tredici 1977, Fyles 1989, Granstom and Schimmel 1993, Livingston and Allessio 1968).Закопанные семена остаются жизнеспособными в почве в течение длительного времени (Fitzsimmons and Burrill 1993).
Прорастание семян
Неизвестно
ЭКОНОМИЧЕСКОЕ ИСПОЛЬЗОВАНИЕ
R. acetosella является плохим кормом для домашнего скота [9] , но выращивается и продается для потребления человеком.
УПРАВЛЕНИЕ
Хороший контроль над Rumex acetosella затруднен, поскольку он размножается и может распространяться как семенами, так и вегетативно. Удаление всходов замедляет или временно препятствует посеву.Эта стратегия также замедляет вегетативное распространение (Фитцсиммонс и Беррилл, 1993). Удаление побегов без удаления корней, по-видимому, стимулирует рост побегов, что приводит к большему процентному покрытию, чем до удаления побегов (Frey et al. 2008).
Возможность восстановления вторженных участков
Воздействие механических средств контроля может привести к образованию сильно нарушенных участков. Это нарушение может быть желательным, а может и нежелательным, в зависимости от природы зараженного растительного сообщества. Если растительное сообщество зависит от нарушения (например, прибрежных дюн), тогда этот метод может быть подходящим.Если, однако, серьезное беспокойство приведет к вымыванию сорняков или иным образом разрушит сообщество, тогда эти методы могут быть неподходящими.
После удаления R. acetosella успех восстановления высок. Нет никаких очевидных химических или физических последствий вторжения.
Профилактика и раннее обнаружение
Как и в случае со всеми нежелательными видами, раннее обнаружение и предотвращение являются наиболее экономически эффективными стратегиями защиты естественной среды обитания от инвазивных видов.
Ручное и механическое управление
Удаление как можно большего количества мелких корней — наиболее эффективный метод уменьшения численности растений. В небольших масштабах это можно эффективно сделать с помощью кирки или лопаты (Frey et al. 2008). После расчистки участка уменьшите повторное прорастание и прорастание семян, применив густую мульчу. (Закат 1998).
Фитцсиммонс и Беррилл (1993) обнаружили, что корни истощаются при повторной культивации в засушливую погоду.
В 2007 году в Президио Сан-Франциско большие пятна плотной р.acetosella . Было протестировано семь методов: 1) использовать пропановую горелку, чтобы увядать листья, 2) использовать мотыгу, чтобы срезать растения чуть ниже поверхности почвы и оставить на месте, 3) не выкапывать растения, вытащить и удалить с участка, 4 ) ручным кирком удалите с участка как можно больше корней и побегов, 5) накройте растительность черной полупроницаемой ландшафтной тканью в течение шести месяцев, 6) удалите 3 дюйма почвы, просейте через сетку и вернуться на участок, 7) удалить 6 дюймов почвы, просеять через сито и вернуться на участок.
Время, необходимое для лечения, сильно варьировалось. Методы удаления корневой биомассы (кирка и копка) снизили процент покрытия R. acetosella . В песках метод извлечения и 6-дюймовая выемка по-прежнему показали снижение процентного покрытия через год после внедрения. В змеевике никакие обработки не показали эффекта через год, но обработка 3-дюймовым копанием уменьшила покрытие до третьего квартала. Результаты показывают, что обработка пламенем является наименее эффективным методом (Frey et al.2008 г.).
Выпас
Щавелевая кислота в составе R. acetosella вызывает дерматит у некоторых животных, и были сообщения о том, что лошади и овцы были отравлены в результате употребления в пищу большого количества этого растения [6] .
R. acetosella пасется овцами и крупным рогатым скотом [9] (Humphrey 1955) и оленями-мулами [9] (Krueger and Donart 1974, Nixon et al. 1970). Тетерев поедают семена овечьего щавеля [9] (Hungerford 1951, Hungerford 1957, Schmidt 1936 и Swenson 1985).
Нет никаких свидетельств того, что выпас используется в качестве метода борьбы.
Предписанный огонь
Не было обнаружено исследований, в которых предписанный огонь использовался в качестве средства борьбы с R. acetosella . Однако имеется некоторая информация о реакции этого вида на огонь. Esser (1995) описывает множество исследований, в которых упоминается R. acetosella и его реакция на огонь [9] (Boyd et al. 1993, Del Tredici 1977, Fyles 1989, Granstrom and Schimmel 1993, Hall 1955, Leege and Godbolt 1985). , MacLean and Wein 1977, Maltby et al.1990 г., Merrill et al. 1980 г., Миноре и др. 1979, Niering and Dreyer 1989). В целом, R. acetosella составляла менее 5% до пожаров и после пожаров. Однако скорость прироста и / или укоренения в целом была выше, чем в контроле.
Наводнение
Неизвестно
Химическая промышленность
50% раствор Rodeo, нанесенный непосредственно на листья R. acetosella в рамках проекта восстановления прибрежных дюн в Президио Сан-Франциско, уменьшил R.acetosella с 42 ± 2 до 7 ± 6 процентов через шесть месяцев (неопубликованные данные).
Университет штата Орегон (1998) рекомендует дикамбу для R. acetosella . Они предлагают 0,5 фунта дикамбы на акр. Они также рекомендуют применять дикамбу, когда красный щавель имеет новую листву поздней осенью, зимой или ранней весной. Они сообщают, что весеннее применение контролирует прорастание весенних всходов лучше, чем ранее применявшиеся опрыскиватели, и что такая обработка предотвратит завязывание семян на выживших растениях (Университет штата Орегон, 1998 г.).
Сансет (1998) рекомендует использовать гербицид, содержащий дикамбу, или обрабатывать пятна глифосатом.
Фитцсиммонс и Беррилл (1993) сообщают, что некоторые гербициды могут избирательно контролировать овечий щавель. Они не перечисляют их, но отсылают читателя к ежегодно издаваемому Справочнику по борьбе с сорняками северо-западного Тихоокеанского региона. Этот документ, теперь называемый PNW Справочник по борьбе с сорняками, можно найти по адресу http://weeds.ippc.orst.edu/pnw/weeds. 6.06.2007 была рекомендована только Дикамба. Рекомендованное время для внесения — «когда красный щавель обретет новую листву, обычно с 15 ноября по 15 марта.Весеннее внесение контролирует прорастание рассады лучше, чем ранее внесенное опрыскивание ». Они также отмечают, что рекомендованная обработка убьет всходы и большинство зрелых растений и предотвратит завязку семян у тех растений, которые действительно выживают (Университет штата Орегон, 2007).
Хотя контроль с помощью гексазинона (триазинового гербицида) показал многообещающие предварительные результаты, будущее исследование показало, что щавель овец вырос и дает большое количество семян [9] (McCully et al.1991).
В Пенсильвании у Solidago spp. и Aster spp. Сообщество, которое было вспахано, дисковым и обработано прометоном, преобладал овечий щавель на 1- и 3-летних участках [9] (Medve 1984).
Биологический контроль
Нет данных о биоконтроле для этого вида.
ПРИМЕРЫ ПРОГРАММ УПРАВЛЕНИЯ Rumex acetosella
Где Rumex acetosella Контроль является регулярной частью восстановительных работ в Президио Сан-Франциско. Повторное удаление несколько раз в год оказалось успешным.
МОНИТОРИНГ
Контроль за распространением этого вида и его раннее обнаружение — ключ к успеху. Кроме того, мониторинг необходим для обнаружения любых изменений в состоянии желаемого сообщества и для того, чтобы увидеть, как сорняк и ваше сообщество реагируют на ваши меры по управлению.
НЕОБХОДИМЫЕ ИССЛЕДОВАНИЯ
- Нехимические методы контроля.
- Сколько биомассы необходимо удалить, чтобы предотвратить цветение?
- Сколько раз и с какой частотой удаление побегов приводит к смерти?
Ссылки
- ↑ ITIS 2007.Интегрированная система таксономической информации. www.itis.gov, дата обращения 26.03.07.
- ↑ IPANE 2007. Атлас инвазивных растений Новой Англии. http://nbii-nin.ciesin.columbia.edu/ipane/, дата обращения 27.03.07. 2,0 2,1 2,2 2,3 2,4 2,5 2,6 2,7 2,8
- ↑ Глисон, Х.А., и А. Кронквист. 1991. Справочник сосудистых растений северо-востока США и прилегающей Канады. 2-е изд. Ботанический сад Нью-Йорка, Нью-Йорк.910 стр.
- ↑ Hickman, J. C. (ed.). 1993. Руководство Джепсона. Беркли, Калифорния: Калифорнийский университет Press. 1400 с.
- ↑ Virginia Tech 2007, Руководство по идентификации сорняков http://www.ppws.vt.edu/scott/weed_id/rumaa.htm, просмотрено 26 марта 2007 г. 5,0 5,1
- ↑ Паркер, К. Ф., 1972. Иллюстрированное руководство по сорнякам Аризоны. Пресса Университета Аризоны, Тусон. Полное онлайн-издание печатной книги. http://www.uapress.arizona.edu/onlinebks/weeds/rsorrel.htm, дата обращения 26.03.07. 6,0 6,1 6,2 6,3 6,4
- ↑ Mabberley, D. J. (1997) The Plant-Book (2-е изд.). Издательство Кембриджского университета, Кембридж.
- ↑ USDA 2007. База данных растений. www.plants.usda.gov, дата обращения 26.03.07. 8,0 8,1 8,2
- ↑ Esser, L. L. 1995. Rumex acetosella. В: Информационная система по пожарным эффектам, [Интернет]. Министерство сельского хозяйства США, Лесная служба, Исследовательская станция Скалистых гор, Лаборатория пожарных наук (производитель).Доступно: http://www.fs.fed.us/database/feis/ Дата обращения: 26.03.07. 9,00 9,01 9,02 9,03 9,04 9,05 9,06 9,07 9,08 9,09 9,10 922 922 922 905 9,17 9,18 9,19 9,20 9,21 9,22 9,23 9.24 9,25 9,26
Другие ресурсы
- APWG 2007. Рабочая группа по чужеродным растениям http://www.nps.gov/plants/alien/list/r.htm, дата обращения 26.03.07.
- Ошмуди, Л. Р., Уолтерс, Р. С. 1988. Восстановление лесов, уничтоженных рассолом. Журнал Американского общества почвоведов. 52: 277-280.
- Бэр, Дж. Э. 1979. Полевые цветы и сорняки Канзаса. Лоуренс, KS: Regents Press Канзаса. 509 с.
- Бойд, Р.С., Вудворд, Р.А., Вальтер, Г. 1993. Воздействие пожара на горном лугу Сьерра-Невада. Калифорнийская рыба и дичь. 70 (3): 115-125.
- Карсон, У. П., Пикетт, С. Т. А. 1990. Роль ресурсов и нарушений в организации сообщества старых полей. Экология. 71 (1): 226-238.
- Кларк Р. А., Феллерс Г. М. 1986. Редкие растения национального побережья Пойнт-Рейес. Tech. Представитель № 22. Дэвис, Калифорния: Калифорнийский университет, Институт экологии, Сан-Франциско, Калифорния: * США. Департамент внутренних дел, Служба национальных парков, Западный регион.117 с.
- Дэвис, Л. Х., Шерман, Р. Дж. 1992. Экологическое исследование редкой Chorizanthe valida (Polygonaceae) на национальном побережье Пойнт-Рейес, Калифорния. Мадроно. 39 (4): 271-280.
- Дельмас М. (ред.) 1993. Fleurs de Vanoise. C.-Y. Шодорель, Эдисуд, Экс-ан-Прованс. 318с.
- Del Tredici, P .. 1977 г. Захороненные семена Comptonia peregrina, сладкого папоротника. Бюллетень Ботанического клуба Торри. 104 (3): 270-275.
- Диттбернер, П. Л., Олсон, М. Р. 1983. База данных информационной сети растений (PIN): Колорадо, Монтана, Северная Дакота, Юта и Вайоминг.FWS / OBS-83/86. Вашингтон, округ Колумбия: Министерство внутренних дел, рыбной ловли и дикой природы США. 786 с.
- Elkhorn Slough 2000. Борьба с сорняками по видам. Национальный эстуарный исследовательский заповедник Элкхорн Слау. http://www.elkhornslough.org/plants/weeds.PDF, дата обращения 06.06.07
- Фидлер П. Л., Лейди Р. А. 1987. Растительные сообщества заповедника Ринг-Маунтин, округ Марин, Калифорния. Мадроно. 34 (3): 173-192.
- Fitzimmons JP и LC Burrill 1993. Красный щавель. Публикация Pacific Northwest Extension.
- Fonda, R. W. 1974. Последовательность лесов в связи с развитием террасы реки в Олимпийском национальном парке, Вашингтон. Экология. 55 (5): 927-942.
- Frey M., J. Soong, J. Feeser и S. Dishy. 2008. Определение методов борьбы с Rumex acetosella в Президио Сан-Франциско (Калифорния). Экологическое восстановление. 26: 2 с.109-111.
- Fyles, J. W. 1989. Популяции банка семян в горных хвойных лесах в центральной Альберте. Канадский журнал ботаники. 67: 274-278.
- Гарднер Р. А. 1958. Почвенно-растительные ассоциации в зоне секвойи и пихты Дугласа в Калифорнии. В: Труды 1-й Североамериканской конференции по лесным почвам, [Дата конференции неизвестна], Ист-Лансинг, Мичиган. Ист-Лансинг, штат Мичиган: Университет штата Мичиган, экспериментальная сельскохозяйственная станция: 86-101.
- GBIF 2007. Глобальный информационный фонд по биоразнообразию. http://www.secretariat.gbif.net/portal/index.jsp, дата обращения 27.03.07,
- Голли, Ф. Б. 1965. Структура и функции сообщества метлочников.Экологические монографии. 35 (1): 113-137.
- Granstrom, A., Schimmel, J. 1993. Тепловое воздействие на семена и корневища некоторых растений бореальных лесов и потенциальная реакция на огонь. Oecologia. 94: 307-313.
- Ассоциация флоры Великих равнин 1986. Флора Великих равнин. Лоуренс, KS: Университетское издательство Канзаса. 1392 с.
- Грин П., Дженсен М. 1991. Последовательность растений в управляемых великих еловых лесах северного Айдахо. В: Харви, А. Э., Нойеншвандер, Л. Ф., составители.Труды — управление и продуктивность лесных почв западных гор, 10-12 апреля 1990 г., Бойсе, штат Иллинойс. Gen. Tech. Реп. INT-280. Огден, Юта: Министерство сельского хозяйства США, Лесная служба, Межгорная исследовательская станция: 232-236.
- Hall, I. V. 1955. Флористические изменения после вырубки и сжигания лесного участка для выращивания черники. Канадский журнал сельскохозяйственных наук. 35: 143-152.
- Halsted, B.D. 1889. Наши худшие сорняки. Ботанический вестник 14 (3): 69-71.
- Голландия, В.Л. 1980. Влияние голубого дуба на кормопроизводство пастбищных угодий в центральной Калифорнии. В: Пламб Т. Р., технический координатор. Материалы симпозиума по экологии, управлению и использованию калифорнийских дубов, 1979, 26-28 июня, Клермонт, Калифорния. Gen. Tech. Реп. PSW-44. Беркли, Калифорния: Министерство сельского хозяйства США, Лесная служба, Тихоокеанская Юго-Западная экспериментальная станция лесов и пастбищ: 314-318.
- Хамфри Р. Р. 1955. Производство кормов на пастбищах Аризоны, IV. Коконино, Навахо, округа Апачи: исследование в условиях ареала.Бюллетень 266. Тусон, Аризона: Университет Аризоны, Экспериментальная сельскохозяйственная станция. 84 с.
- Hungerford, K. E. 1951. Популяции рябчиков и укрытие в северном Айдахо. Сделки, 16-я Североамериканская конференция по дикой природе. [Объем неизвестен]: 216-224.
- Хангерфорд, К. Э. 1957. Оценка кормов для рябчиков для улучшения среды обитания. Транзакции, 22-я Североамериканская конференция по дикой природе. [Объем неизвестен]: 380-395.
- Кирни, Т. Х., Пиблз, Р. Х., Хауэлл, Дж.Т., МакКлинток, Э. 1960. Флора Аризоны. 2-е изд. Беркли, Калифорния: Калифорнийский университет Press. 1085 с.
- Kiltz, B. F. 1930. Многолетние сорняки, распространяющиеся вегетативно. Журнал Американского общества агрономии. 22 (3): 216-234.
- Колб Т. Е., Бауэрсокс Т. В., Маккормик Л. Х. 1990. Влияние интенсивности света на вызванный сорняками стресс саженцев деревьев. Канадский журнал исследований лесного хозяйства. 20: 503-507.
- Крюгер, В. К., Донарт., Г. Б. 1974. Связь почв с сезонным качеством кормов для оленей.Журнал Range Management. 27 (2): 114-117.
- Lackschewitz, K. 1991. Сосудистые растения западно-центральной Монтаны — руководство по идентификации. Gen. Tech. Реп. INT-227. Огден, Юта: Министерство сельского хозяйства США, Лесная служба, Межгорная исследовательская станция. 648 с.
- Leege, T. A., Herman, D. J., Zamora, B. 1981. Влияние выпаса скота на горных лугах в Айдахо. Журнал Range Management. 34 (4): 324-328.
- Leege, T. A., Godbolt, G. 1985. Грибная реакция после предписанного сжигания и посева лосей на ареале в Айдахо.Северо-западная наука. 59 (2): 134-43.
- Линдси, М., Браттон, С. П. 1979. Травянистые лысины Грейт-Смоки-Маунтинс: их история и флора в связи с потенциальным управлением. Управление окружением. 3 (5): 417-430.
- Ливингстон, Р. Б., Алессио, М. Л. 1968. Зарытые жизнеспособные семена в сукцессионных полях и лесных насаждениях, Гарвардский лес, Массачусетс. Бюллетень Ботанического клуба Торри. 95 (1): 58-69.
- MacLean, D. A., Wein, R. W. 1977. Изменения в подлеске с увеличением возраста древостоя в лесах Нью-Брансуика: видовой состав, покров, биомасса и питательные вещества.Канадский журнал ботаники. 55: 2818-2831.
- Maltby, E., Legg, C.J., Proctor, M.C.F. 1990. Экология сильного пожара в болотах Северного Йорка: последствия пожаров 1976 года и последующее развитие поверхности и растительности. Журнал экологии. 78 (2): 490-518.
- Марк, А. Ф. 1958. Экология лысины южных Аппалачей. Экологические монографии. 28 (4): 293-336.
- Макбрайд, Дж. Р., Норберг, Э., Ченг, С., Мосаддык, А. 1991. Посадка рассады прибрежного живого дуба в связи с сохранением семян сойкой.В: Стэндифорд, Р. Б., технический координатор. Материалы симпозиума по дубовым лесам и управлению пастбищами лиственных пород, 31 октября — 2 ноября 1990 г., Дэвис, Калифорния. Gen. Tech. Реп. PSW-126. Беркли, Калифорния: Министерство сельского хозяйства США, Лесная служба, Тихоокеанская юго-западная исследовательская станция: 143-148.
- Маккалли, К. В., Сэмпсон, М. Г., Уотсон, А. К. 1991. Обследование сорняков на полях низкорослой голубики (Vaccinium angustifolium) в Новой Шотландии (Канада). Наука о сорняках. 39 (2): 180-185.
- Медве, Р.J. 1984. Микориза первых видов в нарушенных экосистемах западной Пенсильвании. Американский журнал ботаники. 71 (6): 787-794.
- Меррилл, Э. Х., Мэйленд, Х. Ф., Пик, Дж. М. 1980. Влияние осеннего лесного пожара на травянистую растительность на ксерических участках в пустыне Селуэй-Биттеррут, штат Айдахо. Журнал Range Management. 33 (5): 363-367.
- Миноре, Д., Смарт, А. В., Дубрасич, М. Е. 1979. Экология Гекльберри и исследования в области управления на северо-западе Тихого океана. Gen. Tech.Представитель PNW-93. Портленд, штат Орегон: Министерство сельского хозяйства, лесной службы США, Тихоокеанская Северо-Западная экспериментальная станция лесов и пастбищ. 50 шт.
- Niering, W. A., Dreyer, G. D. 1989. Влияние предписанного сжигания на Andropogon scoparius на постсельскохозяйственных лугах в Коннектикуте. Американский натуралист из Мидленда. 122: 88-102.
- Никсон, К. М., Макклейн, М. В., Рассел, К. Р. 1970. Пищевые привычки и характеристики ареала оленей в Огайо. Журнал управления дикой природой. 34 (4): 870-886.
- Государственный университет Орегона 1998.Программы по борьбе с сорняками в Орегоне
http://cropandsoil.oregonstate.edu/weeds/Red_sorrel/control.html. Последнее обновление: 7 апреля 1998 г., дата обращения 26.03.07.
- Университет штата Орегон 2007 PNW Справочник по борьбе с сорняками. http://weeds.ippc.orst.edu/pnw/weeds. по состоянию на 06.06.2007.
- Рэдфорд, А. Э., Алес, Х. Э., Белл, К. Ричи. 1968. Справочник сосудистой флоры Каролины. Чапел-Хилл, Северная Каролина: Издательство Университета Северной Каролины. 1183 с.
- Рэнд, Э. Л.1890. Некоторые дальнейшие заметки о флоре озер Рэнджли. Бюллетень Ботанического клуба Торри 17 (2): 32-34.
- Шмидт, Ф. Дж. У. 1936. Зимний корм острохвостого тетерева и перистого тетерева в Висконсине. Бюллетень Вильсона. Сентябрь: 186-203.
- Сугихара, Н. Г., Рид, Л. Дж., Ленихан, Дж. М. 1987. Растительность дубовых лесов Лысых холмов, Национальный парк Редвуд, Калифорния. Мадроно. 34 (3): 193-208.
- Закат для редакторов книг и журналов. 1998. Решение проблем «Западный сад» Закат.Сансет Букс, Инк., Менло-Парк, Калифорния.
- Стрейт, Р. А., Джексон, М. Т. 1986. Экологический анализ растительных сообществ природного заповедника Литл-Блюстем-Прери: предварительное сжигание по сравнению с последующим сжиганием. Слушания, Академия наук Индианы. 95: 447-452.
- Strausbaugh, P. D., Core, E. L. 1977. Флора Западной Вирджинии. 2-е изд. Моргантаун, Западная Вирджиния: Seneca Books, Inc. 1079 стр.
- Свенсон, Дж. Э. 1985. Сезонное использование среды обитания острохвостым тетеревом, Tympanuchus phasianellus, в прериях с разнотравьем в Монтане.Канадский естествоиспытатель. 99 (1): 40-46.
- Тильман Д. 1988. Динамика и структура растительных сообществ. Монографии по популяционной биологии 26. Princeton, NJ: Princeton University Press. 360 с.
- Уолтерс Р. С., Ошмуди Л. Р. 1989. Восстановление растительности на лесном массиве твердых пород, оголенном рассолом. Ландшафт и градостроительство. 17: 127-133.
- Уивер Т., Лихтхарт Дж., Густафсон Д. 1990. Экзотическое вторжение в лесную растительность, Северные Скалистые горы, США.В: Шмидт, У. К., Макдональд, К. Дж., Составители. Труды симпозиума по экосистемам белой сосны: экология и управление высокогорным ресурсом, 1989 г., 29-31 марта, Bozeman, MT. Gen. Tech. Представитель INT-270. Огден, UT:. Wofford, B. E. 1989. * Путеводитель по сосудистым растениям Голубого хребта. Афины, Джорджия: Издательство Университета Джорджии. 384 с. Министерство сельского хозяйства США, Лесная служба, Межгорная исследовательская станция: 208-213.
- Wofford, B. E. 1989. Руководство по сосудистым растениям Голубого хребта.Афины, Джорджия: Издательство Университета Джорджии. 384 с.
- Циммерман, Г. Т., Нойеншвандер, Л. Ф. 1984. Влияние выпаса скота на структуру сообщества, интенсивность пожаров и частоту пожаров в пределах типа среды обитания пихты Дугласовой / девятикорной. * Журнал Range Management. 37 (2): 104-110.
|
Польза для здоровья, использование, побочные эффекты, дозировка и взаимодействие
Бхакуни, Д.С., Биттнер, М., Мартикорена, К., Сильва, М., Велдт, Э., Хоенизен, М. Скрининг чилийских растений на противораковую активность. И. Ллойдия. 1976; 39 (4): 225-243. Просмотреть аннотацию.
Чоу С., Хван Б., Ким М. и др. Химические компоненты Rumex acetellosa L. Korean J Pharmacog 1998; 29: 209-216.
Дорнбергер К. и Лич Х. [Скрининг антимикробных и предполагаемых канцеростатических метаболитов растений (авторский перевод)]. Pharmazie 1982; 37 (3): 215-221. Просмотреть аннотацию.
Эрнст, Э., Марц, Р. У., и Сидер, К. [Острый бронхит: эффективность Синупрета. Сравнительное исследование с обычными отхаркивающими средствами у 3187 пациентов]. Fortschr.Med 4-20-1997; 115 (11): 52-53. Просмотреть аннотацию.
Фарре М., Ксиргу Дж., Сальгадо А., Перакаула Р., Рейг Р. и Санз П. Смертельное отравление щавелевой кислотой из щавелевого супа. Ланцет 12-23-1989; 2 (8678-8679): 1524. Просмотреть аннотацию.
Гняздовска Б., Дорошевска Г., Дорошевский В. Гиперчувствительность к аллергенам пыльцы сорных растений в районе Бигдощи.Пневмонол.Алергол.Пол. 1993; 61 (7-8): 367-372. Просмотреть аннотацию.
Карн Х. и Мур MJ. Использование лечебного средства на травах ESSIAC в амбулаторных онкологических условиях. Proc Annu Meet Am Soc Clin Oncol 1997; 16: A245.
Ли, Н. Дж., Чой, Дж. Х., Ку, Б. С., Рю, С. Ю., Хан, Ю. Х., Ли, С. И. и Ли, Д. У. Антимутагенность и цитотоксичность компонентов из надземных частей Rumex acetosa. Биол Фарм Булл. 2005; 28 (11): 2158-2161. Просмотреть аннотацию.
Локок RA. Фитотерапия: Essiac.Can Pharm J 1997; 130 (февраль): 18-19, 51.
Melzer, J., Saller, R., Schapowal, A., and Brignoli, R. Систематический обзор клинических данных с BNO-101 (Sinupret) при лечении гайморита. Forsch Komplement.Med (2006) 2006; 13 (2): 78-87. Просмотреть аннотацию.
Neubauer N и Marz RW. Плацебо-контролируемое рандомизированное двойное слепое клиническое исследование таблеток Синупрет, покрытых сахарной оболочкой, на основе терапии антибиотиками и противоотечными назальными каплями при остром синусите. Фитомедицина 1994; 1: 177-181.
Ричардсон, М.А. Исследования методов дополнительной / альтернативной медицины в онкологии: многообещающие, но сложные. J Clin Oncol. 1999; 17 (11 доп.): 38-43. Просмотреть аннотацию.
Ричштейн А. и Манн В. Лечение хронического синусита Синупретом. Ther.Ggw. 1980; 119 (9): 1055-1060. Просмотреть аннотацию.
Sanz, P. и Reig, R. Клинические и патологические данные при летальном оксалозе растений. Обзор. Am J Forensic Med Pathol. 1992; 13 (4): 342-345. Просмотреть аннотацию.
Тай, Дж.и Cheung, S. Исследования in vitro культуры FlorEssence на линиях опухолевых клеток человека. Phytother Res 2005; 19 (2): 107-112. Просмотреть аннотацию.
Тарасова Г.Д. Применение синупрета в лечении острого синусита у детей. Вестн.Оториноларингол. 2001; (2): 46-48. Просмотреть аннотацию.
Офис оценки технологий Конгресса США. Эссиак. Типография правительства США, Вашингтон, округ Колумбия: 1990.
Ямамото А. Эссиак. Кан Дж. Хосп Фарм 1988; 41 (3): 158.
Измаил, К., Визель, А., Марц, Р. В., и Кайссер-Люфт, А. Эпиднадзорное исследование Sinupret в сравнении с данными реестра рождений Майнца. Arch Gynecol. Obstet 2003; 267 (4): 196-201. Просмотреть аннотацию.
Ито Х. Влияние противоопухолевых агентов из различных природных источников на систему метаболизма лекарств, фагоцитарную активность и систему комплемента у мышей, несущих саркому 180. Jpn J Pharmacol 1986; 40: 435-43. Просмотреть аннотацию.
Джабер Р. Респираторные и аллергические заболевания: от инфекций верхних дыхательных путей до астмы.Prim Care 2002; 29: 231-61. Просмотреть аннотацию.
Каэги Э. Нетрадиционные методы лечения рака: 1. Essiac. Целевая группа по альтернативным методам лечения Канадской инициативы по исследованию рака груди. CMAJ 1998; 158: 897-902. Просмотреть аннотацию.
Marz RW, Ismail C, Popp MA. Профиль действия и эффективность комбинированного травяного препарата для лечения синусита. Wien Med Wochenschr 1999; 149: 202-8.