
ООО «ФАКТОР СВЕТА», реквизиты, адрес, телефон, контакты, отзывы, вакансии 2021
Все компании • Розничная торговля, кроме торговли автотранспортными средствами и мотоциклами, ремонт бытовых изделий и предметов личного пользования • ООО «ФАКТОР СВЕТА»ООО «ФАКТОР СВЕТА»: адрес, телефон, факс, email, сайт, график работы
ОБЩЕСТВО С ОГРАНИЧЕННОЙ ОТВЕТСТВЕННОСТЬЮ «ФАКТОР СВЕТА»
Регион: Волгоградская область Волгоград
Адрес: 400117, г. ВОЛГОГРАД, ул. ИМ. ЗЕМЛЯЧКИ, 62А, кв. 81
Телефон: 8442-22-76-36, 8-961-690-08-62
Факс: нет данных
E-mail: нет данных
Сайт: нет данных
Генеральный директор / ответственное лицо / владелец ООО «ФАКТОР СВЕТА»: нет данных
График работы:
Подробная информация об ООО «ФАКТОР СВЕТА»: бухгалтерия, баланс. Скачать банковские реквизиты, тендеры, кредитная история, налоги ООО «ФАКТОР СВЕТА».
Скачать банковские реквизиты, тендеры, кредитная история, налоги ООО «ФАКТОР СВЕТА».
ОГРН: 1123443001494
ИНН: 3443114901
КПП:
ОКПО: 9551836
ОКАТО: 18401365000
Получить выписку из ЕГРЮЛ об ООО «ФАКТОР СВЕТА»
Общества с ограниченной ответственностью
Частная собственность
Виды деятельности по ОКВЭД:
Розничная торговля, кроме торговли автотранспортными средствами и мотоциклами, ремонт бытовых изделий и предметов личного пользования
Розничная торговля светильниками
Работа в
ООО «ФАКТОР СВЕТА» вакансии, практика, стажировка, карьераНа данный момент открытых вакансий нет. Возможно вас заинтересуют вакансии в других компаниях:
Возможно вас заинтересуют вакансии в других компаниях:
Менеджер по обслуживанию (г.Екатеринбург, ул.Щербакова, 20)
Водитель-экспедитор
Оператор ПК
Машинист грунтового катка
Торговый представитель, менеджер по продажам
Мастер маникюра, педикюра в современный салон г.Омска.
Повар г/ц
Водитель на самосвал
Продавец-грузчик магазина Магнит
Фасовщик корма
Продавец-логист — Московка-2
Продавец-консультант в магазин женской одежды
Отзывы об ООО «ФАКТОР СВЕТА»
Отзывы об ООО «ФАКТОР СВЕТА»
Карта проезда к ООО «ФАКТОР СВЕТА» где находится
Также смотрите компании и организации с похожим видом деятельности, как у ООО «ФАКТОР СВЕТА»: ООО «ВИГ» | ООО «ЭНЕРГИЯ» | ООО «ЛИДЕК» | ООО ТОРГОВЫЙ РЯД НН | ООО «ТО ГЕРМЕС»
Полное название: ОБЩЕСТВО С ОГРАНИЧЕННОЙ ОТВЕТСТВЕННОСТЬЮ «ФАКТОР СВЕТА», ОГРН: 1123443001494, ИНН: 3443114901. Регион: Волгоградская область, г. Волгоград. Фирма ООО «ФАКТОР СВЕТА» расположена по адресу: 400117, г. ВОЛГОГРАД, ул. ИМ. ЗЕМЛЯЧКИ, 62А, кв. 81. Основной вид деятельности: «Розничная торговля, кроме торговли автотранспортными средствами и мотоциклами, ремонт бытовых изделий и предметов личного пользования / Розничная торговля мебелью и товарами для дома / Розничная торговля светильниками».
Регион: Волгоградская область, г. Волгоград. Фирма ООО «ФАКТОР СВЕТА» расположена по адресу: 400117, г. ВОЛГОГРАД, ул. ИМ. ЗЕМЛЯЧКИ, 62А, кв. 81. Основной вид деятельности: «Розничная торговля, кроме торговли автотранспортными средствами и мотоциклами, ремонт бытовых изделий и предметов личного пользования / Розничная торговля мебелью и товарами для дома / Розничная торговля светильниками».
Второй фактор: как подключить и отключить
Чтобы дополнительно защитить аккаунт от злоумышленников, любой пользователь Wargaming.net всегда может подключить второй фактор защиты аккаунта. Мобильный аутентификатор — приложение на вашем мобильном устройстве, которое позволяет создавать одноразовые пароли для входа в Личный кабинет и восстановления доступа к аккаунту. Это наилучший вариант защиты аккаунта.
Чтобы его подключить, вам понадобится установить на своё мобильное устройство специальное приложение — Google Authenticator или Microsoft Authenticator.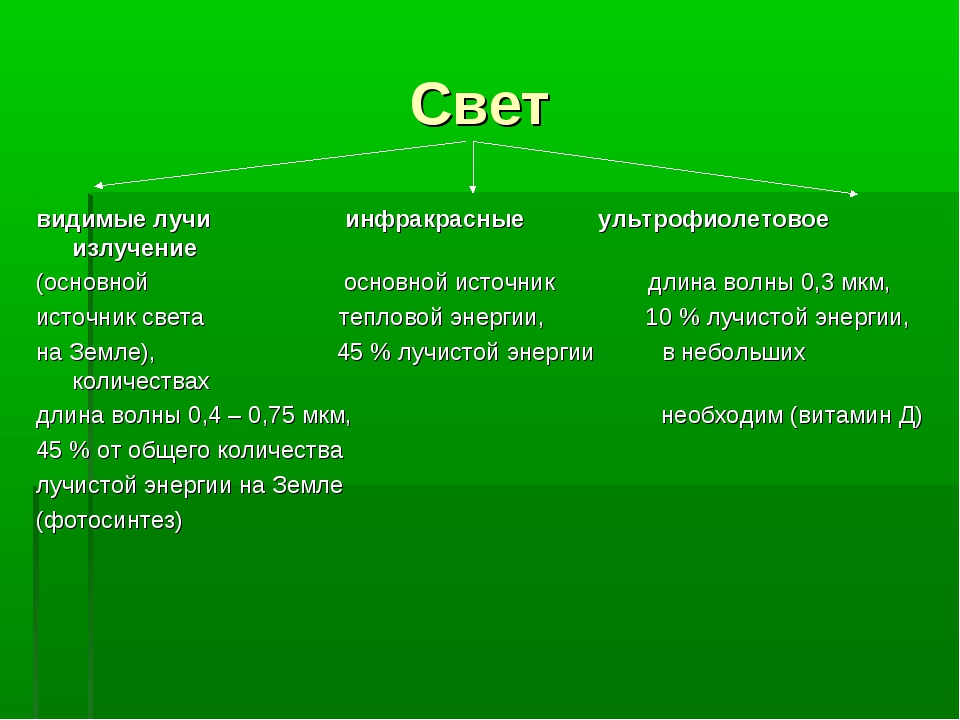
Подключив второй фактор защиты, при входе в Личный кабинет и других важных операциях (восстановлении аккаунта, создании резервных кодов) вы будете дополнительно использовать специальный код аутентификации, сгенерированный этим приложением.
Обязательно сохраните секретный ключ от мобильного аутентификатора. Он понадобится вам, если вы потеряете телефон и захотите восстановить доступ к аккаунту.
Подключение второго фактора защиты к игровому аккаунту — бесплатная операция. Доступна только для устройств с системами iOS и Android.
Как подключить, если аккаунт не привязан к телефону
Как подключить, если аккаунт привязан к телефону
Как установить Google Authenticator
Приложение Google Authenticator используется для создания кодов двухэтапной аутентификации в вашем телефоне.
Двухэтапная аутентификация обеспечивает более надёжную защиту учётной записи: чтобы войти в аккаунт, потребуется не только пароль, но и код подтверждения.
Активировать приложение Google Authenticator можно только после включения двухэтапной аутентификации.
Приложение Google Authenticator работает на устройствах iPhone, iPod Touch и iPad с iOS 5.0 и более поздних версий. Настроить приложение с помощью QR-кода можно только на iPhone 3G или более новых моделях.
Приложение Google Authenticator работает на устройствах Android версии 2.1 и более поздних.
Установка приложения
- В App Store или Google Play введите в строке поиска Google Authenticator. Скачайте и установите приложение.
- Зайдите в приложение и нажмите Приступить к настройке.
- Привяжите мобильное устройство к аккаунту одним из способов:
- Способ 1 — с помощью QR-кода:
Нажмите «Сканировать штрихкод», а затем наведите камеру на QR-код, отображённый на экране компьютера. - Способ 2 — с помощью секретного ключа:
В поле «Ввод вручную» укажите адрес электронной почты. В поле «Ключ» введите секретный ключ и нажмите на галочку вверху страницы.
В поле «Ключ» введите секретный ключ и нажмите на галочку вверху страницы.
- Способ 1 — с помощью QR-кода:
- Сохраните свой секретный ключ в надёжном месте, чтобы в случае утери доступа ко второму фактору защиты вы могли самостоятельно его восстановить.
- Аккаунт успешно привязан.
- Подключив второй фактор защиты аккаунта, не удаляйте приложение. Если вам понадобится удалить приложение, перед этим обязательно отключите аутентификацию.
- Если вы всё-таки удалили приложение Google Authenticator, не отключив перед этим аутентификацию, воспользуйтесь инструкцией для восстановления доступа.
 В поле «Ключ» введите секретный ключ и нажмите на галочку вверху страницы.
В поле «Ключ» введите секретный ключ и нажмите на галочку вверху страницы.Как установить Microsoft Authenticator
Как отключить второй фактор защиты
✅ ООО «ФАКТОР СВЕТА», 🏙 Москва (OГРН 5167746402676, ИНН 7707376240, КПП 770901001) — 📄 реквизиты, 📞 контакты, ⭐ рейтинг
Последствия пандемии
В полной версии сервиса доступна вся информация по компаниям, которых коснулись последствия пандемии коронавируса: данные об ограничениях работы и о программе помощи от государства тем отраслям, которые испытывают падение спроса
Получить доступКраткая справка
ООО «ФАКТОР СВЕТА» было зарегистрировано 02 декабря 2016 (существовало 2 года) под
ИНН 7707376240 и
ОГРН 5167746402676. Юридический адрес 109029, Москва, Калитниковская Б. улица, дом 42.
Руководитель КАМКИНА ЮЛИЯ БОРИСОВНА.
Основной вид деятельности ООО «ФАКТОР СВЕТА»: 43.21 Производство электромонтажных работ.
Телефон, адрес электронной почты, адрес официального сайта и другие контактные данные ООО «ФАКТОР СВЕТА» отсутствуют в ЕГРЮЛ.
Ликвидировано 23 апреля 2019.
Юридический адрес 109029, Москва, Калитниковская Б. улица, дом 42.
Руководитель КАМКИНА ЮЛИЯ БОРИСОВНА.
Основной вид деятельности ООО «ФАКТОР СВЕТА»: 43.21 Производство электромонтажных работ.
Телефон, адрес электронной почты, адрес официального сайта и другие контактные данные ООО «ФАКТОР СВЕТА» отсутствуют в ЕГРЮЛ.
Ликвидировано 23 апреля 2019.
Информация на сайте предоставлена из официальных открытых государственных источников.
Контакты ООО «ФАКТОР СВЕТА»
Основной адрес
109029, Россия, Москва, Калитниковская Б. улица, дом 42
Зарегистрирован 02 декабря 2016
Перейти ко всем адресам
Телефоны
8 (495). .. показать
.. показать Электронная почта
—Роль фактора света в жизни растений
Свет, несомненно, является главным фактором жизнедеятельности растений.
Без энергии солнечного света невозможен процесс фотосинтеза, процесс аккумуляции (накопления) солнечной энергии на земле.
Следует различать прямой и рассеянный свет. Рассеянный свет полезен для растений — в нем больше сине-фиолетовых и оранжево-красных лучей (50—60 %). В прямом свете этих лучей только около 37 %. Для нормального развития растений важно правильное соотношение прямого и рассеянного света. Это соотношение может быть различным, так как оно зависит от географического положения участка, высоты солнца, времени суток, содержания в воздухе паров воды, пыли и т. д. В северных районах страны больше рассеянного света, в южных — прямого.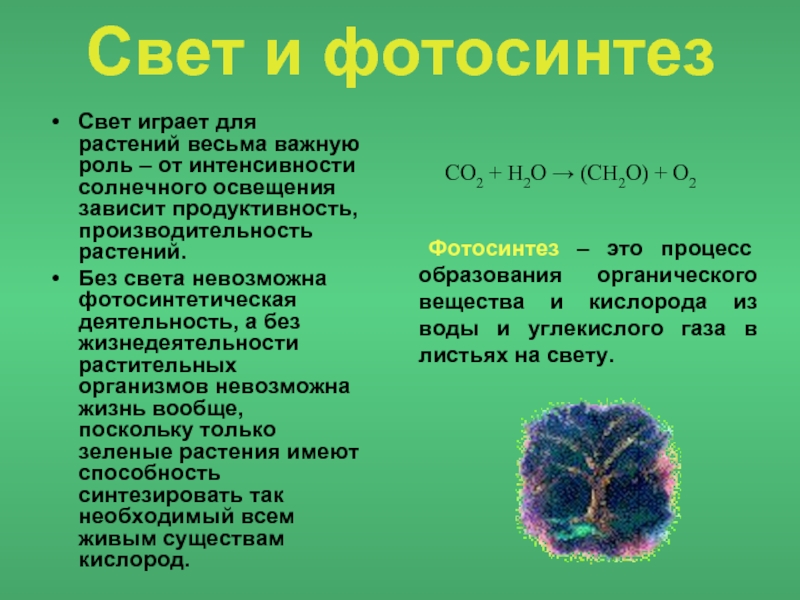
Количество суммарной (прямой и рассеянной) солнечной радиации, поступающей на поверхность земли, увеличивается от полюсов к экватору. Так, в Ленинграде ее приток за год примерно в два раза меньше, чем в Крыму. Но если сравнить количество поступающей суммарной радиации только за летние месяцы, то разница составит менее 30 %. По расчетам А. П. Шенникова, при усвоении растениями только одного процента поступаемой световой энергии можно даже на Шпицбергене с одного гектара получать ежегодно около пяти тонн сухого органического вещества, но из-за отсутствия необходимого количества тепла осуществить это невозможно.
Важное значение для развития растений имеет длина дня. Это особенно относится к северным районам страны. Во-первых, при длинном дне и короткой ночи уменьшается вероятность наступления весенних заморозков. За короткую ночь земная поверхность не успевает сильно охладиться в результате эффективного излучения.
Во-вторых, в весенне-летний период фотосинтез возможен почти круглосуточно в районах вблизи Полярного круга, а выше его — в течение всех суток. Доказано, что при гражданских сумерках (что характерно для широты близ Полярного круга) фотосинтез не прекращается.
В-третьих, при сочетании длинного дня с невысокими температурами воздуха, особенно при низком стоянии солнца, соотношение фотосинтеза и дыхания смещается в сторону «экономного» расходывания ассимилятов на дыхание. Это способствует интенсивному накоплению биомассы растениями. Так, урожайность картофеля, посаженного в Заполярье в первой половине июня, уже к началу августа достигает 15—20 т/га, а к концу августа — 25—30 т/га и более.
В защищенном грунте на севере Европейской части страны и Урало-Сибирского региона при естественном освещении овощные растения можно успешно выращивать в течение полугода — с 21 марта по 23 сентября, т. е. в период между весенним и осенним равноденствием.
Влияние света следует рассматривать во взаимосвязи с такими факторами среды, как температура, обеспеченность водой, кислородом, углекислотой, элементами минерального питания. Эти факторы все время меняются и находятся в постоянной зависимости от времени суток, облачности, силы ветра и т. д. При этом в определенные моменты один фактор может быть в минимуме, другой — в максимуме, а третий — в оптимуме. В какие-то периоды все факторы будут в оптимуме или близки к нему. В такие периоды растения растут и развиваются очень быстро.
Эти факторы все время меняются и находятся в постоянной зависимости от времени суток, облачности, силы ветра и т. д. При этом в определенные моменты один фактор может быть в минимуме, другой — в максимуме, а третий — в оптимуме. В какие-то периоды все факторы будут в оптимуме или близки к нему. В такие периоды растения растут и развиваются очень быстро.
Установлено, что для формирования репродуктивных органов (органов воспроизведения) растения нуждаются в большей облученности, чем для формирования вегетативных органов (органов для поддержания жизни). Вот почему выгоночные растения и растения, формирующие урожай за счет вегетативных органов (лук, капуста, салат и др.), менее требовательны к свету и температуре, чем растения, формирующие урожай за счет репродуктивных органов (томат, огурец, перец и др.).
Рост и развитие растений в северных районах при обеспеченности другими факторами жизни находятся в прямой зависимости от светового насыщения, которое в условиях Севера, как правило, не является избыточным. Только при освещенности выше 70—80 тысяч люкс свет может в отдельных случаях повлиять отрицательно, вызывая у растений перегрев и ожоги. Вредны резкие перепады освещенности от низкой к высокой, и наоборот.
Только при освещенности выше 70—80 тысяч люкс свет может в отдельных случаях повлиять отрицательно, вызывая у растений перегрев и ожоги. Вредны резкие перепады освещенности от низкой к высокой, и наоборот.
Свет оказывает большое влияние на развитие корневой системы: при интенсивном освещении она развивается быстрее, чем надземные органы. Если снижается интенсивность света, то следует обязательно снижать температуру почвы и воздуха, особенно при выращивании рассады. В противном случае рассада очень быстро вытягивается и теряет свои качества. Весьма требовательны к свету плодовые и ягодные культуры, они плохо переносят даже частичное затенение; при затенении снижается не только урожайность плодов и ягод, но и зимостойкость, что крайне нежелательно, особенно в условиях Севера.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Свет как экологический фактор
Свет наиболее важен для растений. Он поставляет энергию для фотосинтеза. Фотосинтез фактически активирует процессы в большинстве экосистем. Солнце является источником света. В рамках солнечного спектра выделяют следующие варианты.
Фотосинтез фактически активирует процессы в большинстве экосистем. Солнце является источником света. В рамках солнечного спектра выделяют следующие варианты.
1. Коротковолновое ультрафиолетовое излучение, источник мутагенеза. Это излучение мало проходит на Землю, отражается озоновым слоем, и его, например, могут различать насекомые.
2. Видимая часть светового спектра преобладает и обеспечивает эффективную работу органов зрения.
3. Инфракрасные, или тепловые волны имеют максимальную длину волны, могут использоваться некоторыми бактериями для фотосинтеза.
Выделяют три основные группы растений по отношению к свету
1. Светолюбивые растения (обитают на открытых местах).
Светолюбивые растения (обитают на открытых местах).
2. Теневыносливые (деревья леса).
3. Тенелюбивые (лесные травы, мхи).
Среди гетеротрофов для некоторых губителен свет (бактерии), для большинства он необходим для ориентации в пространстве. Длина светового дня может изменяться, поэтому существуют суточные ритмы. В соответствии с ними организмы синхронизируют свою активность. Растения цветут строго при определённой продолжительности дня, что определило их разделение на две группы.
1. Короткодневные — цветут при коротком дне.
2. Длиннодневные — для цветения им требуется световой день более 14 часов.
Признаки светолюбивых растений (гелиофитов)
1. Часто укороченные побеги, сильно рассечённые листья, развитая механическая и запасающая ткань (василёк луговой, ковыль, полынь).
Часто укороченные побеги, сильно рассечённые листья, развитая механическая и запасающая ткань (василёк луговой, ковыль, полынь).
2. Цветки поворачиваются к свету. Они же любят свет!
3. У листьев растений открытых мест светлая окраска, т. к. содержат меньше хлорофилла.
Признаки теневыносливых растений (факультативные гелиофиты)
1. Растут на свету, но выдерживают и его недостаток.
2. В глубине кроны — теневые листья, на периферии — световые.
3. Имеют более тёмную окраску, так как содержат больше хлорофилла.
4. Цветки у теневыносливых растений часто белые для привлечения насекомых в тени.
5. Хлоропласты крупные.
6. Примеры — сирень, липа сердцелистная, земляника лесная.
Тенелюбивые растения (сциофиты)
1. Растения нижних ярусов лесов, глубоких водоёмов.
2. Используют лишь 0,1—0,2 процента падающего света, плохо переносят освещение прямыми солнечными лучами.
3. Вытянутые побеги, листья тёмно-зелёные, крупные и тонкие, в них плохо развита механическая ткань.
4. К ним относятся мхи, плауны, кислица обыкновенная, красные и бурые водоросли.
Животные по отношению к свету делятся на
1. Дневные (белка европейская).
2. Сумеречные (майский жук).
3. Ночные (серый ушан).
 Восприятие света и цвета некоторыми животными» src=»//www.youtube.com/embed/-kVePSp1qD4?feature=oembed» frameborder=»0″ allow=»accelerometer; autoplay; clipboard-write; encrypted-media; gyroscope; picture-in-picture» allowfullscreen=»»>
Восприятие света и цвета некоторыми животными» src=»//www.youtube.com/embed/-kVePSp1qD4?feature=oembed» frameborder=»0″ allow=»accelerometer; autoplay; clipboard-write; encrypted-media; gyroscope; picture-in-picture» allowfullscreen=»»>
Каждый процент света зимой обеспечивает чистую прибыль : ReduSystems
AntiReflect
Часть света, падающего под углом, отражается от тепличной крыши. Чем меньше угол падения, тем больше света будет отражаться, а зимой угол падения всегда будет меньше. Отражённый свет не несёт никакой пользы для культуры. В этом сезоне Mardenkro вместе с голландскими растениеводами испытывает новое покрытие AntiReflect, которое повышает светопропускаемость теплицы на 3%. Первых пользователей мы поддерживаем, помогая производить измерения светопропускаемости.
Конденсат
Конденсат на внутренней стороне крыши также может привести к значительным потерям света. Зимой этот фактор очень важен, поскольку в это время крыша почти постоянно влажная. Тип конденсата также определяет пропускание света. Большие капли поглощают целых 5-8% света. Равномерно распределенная водная плёнка пропускает больше света. Новое покрытие AntiCondens для стекла действует как смачивающий агент и создает пленку воды поверх стекла. Первоначальный опыт его применения на практике был положительным: измерения, проведенные в теплице с томатами в мае 2017 года, показали, что проникновение света с использованием AntiCondens было на 6-7% выше, чем без него.
Тип конденсата также определяет пропускание света. Большие капли поглощают целых 5-8% света. Равномерно распределенная водная плёнка пропускает больше света. Новое покрытие AntiCondens для стекла действует как смачивающий агент и создает пленку воды поверх стекла. Первоначальный опыт его применения на практике был положительным: измерения, проведенные в теплице с томатами в мае 2017 года, показали, что проникновение света с использованием AntiCondens было на 6-7% выше, чем без него.
Свет после попадания внутрь теплицы должен сразу поглощаться растениями, а не полом или элементами конструкции. Поэтому внутренняя часть теплицы должна быть максимально отражающей: белое напольное покрытие и порошковое покрытие на всех конструкциях. Порошковое покрытие отражает около 80% света. Можно использовать PipePaint от Mardenkro для того, чтобы повысить светоотражаемость отопительных труб и предотвратить коррозию.
Теплица с зимним типом освещения
Отделение тепличного растениеводства в Университете Wageningen в течение нескольких лет работает над созданием теплицы, оптимизированной для зимнего использования: Теплица с зимним типом освещения. Первый шаг состоял в том, чтобы определить, какие факторы оказывают наибольшее влияние (главным образом, из тех, что описаны выше). Модель этой теплицы начала работать в феврале прошлого года в Блейсвейке. Теплица построена из высокопрозрачного диффузного стекла, которое специально протравлено, чтобы сделать его антибликовым. Высокопрозрачный инновационный экран устанавливается в форме W для максимальной передачи света в закрытом состоянии. Тепличная конструкция обработана специальным порошковым покрытием, которое отражает 87% света.
Первый шаг состоял в том, чтобы определить, какие факторы оказывают наибольшее влияние (главным образом, из тех, что описаны выше). Модель этой теплицы начала работать в феврале прошлого года в Блейсвейке. Теплица построена из высокопрозрачного диффузного стекла, которое специально протравлено, чтобы сделать его антибликовым. Высокопрозрачный инновационный экран устанавливается в форме W для максимальной передачи света в закрытом состоянии. Тепличная конструкция обработана специальным порошковым покрытием, которое отражает 87% света.
Расчеты модели, выполненные до строительства, показали, что потенциальное усиление света может достигать 10,5%, и измерения в теплице подтвердили это. В настоящий момент в этой теплице выращивают огурцы. Выращивание длится до начала декабря. В текущий момент производительность приближается к 110 кг / м².
Свет как абиотический фактор — Docsity
Содержание Введение 1.Абиотические факторы 2.Свет как абиотический фактор 3.Заключение 4.Список использованной литературы Введение Пространство, в котором живут организмы, и все то, что их окружает в этом пространстве, есть окружающая среда. Среда — это совокупность элементов, которые способны оказывать прямое или косвенное воздействие на организмы. Элементы окружающей среды, оказывающие влияние на живые организмы» называются экологическими факторами. Они подразделяются на абиотические, биотические и антропогенные. К числу абиотических факторов относятся элементы неживой природы: свет, температура, влажность, осадки, ветер, атмосферное давление, радиационный фон, химический состав атмосферы, воды, почвы и т. п. Биотическими факторами являются живые организмы (бактерии, грибы, растения, животные), вступающие во взаимодействие с данным организмом. К антропогенным факторам относятся особенности среды, обусловленные трудовой деятельностью человека. По мере роста народонаселения и технической вооруженности человечества удельный вес антропогенных факторов постоянно возрастает. Следует учитывать, что на отдельные организмы и их популяции одновременно воздействуют многие факторы, создающие определенный комплекс условий, в котором могут обитать те или иные организмы.
Среда — это совокупность элементов, которые способны оказывать прямое или косвенное воздействие на организмы. Элементы окружающей среды, оказывающие влияние на живые организмы» называются экологическими факторами. Они подразделяются на абиотические, биотические и антропогенные. К числу абиотических факторов относятся элементы неживой природы: свет, температура, влажность, осадки, ветер, атмосферное давление, радиационный фон, химический состав атмосферы, воды, почвы и т. п. Биотическими факторами являются живые организмы (бактерии, грибы, растения, животные), вступающие во взаимодействие с данным организмом. К антропогенным факторам относятся особенности среды, обусловленные трудовой деятельностью человека. По мере роста народонаселения и технической вооруженности человечества удельный вес антропогенных факторов постоянно возрастает. Следует учитывать, что на отдельные организмы и их популяции одновременно воздействуют многие факторы, создающие определенный комплекс условий, в котором могут обитать те или иные организмы. Одни факторы могут усиливать или ослаблять действие других факторов. Например, при оптимальной температуре повышается выносливость организмов к недостатку влаги и пищи; в свою очередь обилие пищи увеличивает устойчивость организмов к неблагоприятным климатическим обладать цветовым зрением, а у растений цветки имеют яркую окраску для привлечения насекомых-опылителей. Инфракрасные лучи с длиной волны более 0,750 мкм глаз человека не воспринимает, но они являются источником тепловой энергии (45% лучистой энергии). Эти лучи поглощаются тканями животных и растений, вследствие чего ткани нагреваются. У растений важнейшая функция инфракрасных лучей состоит в осуществлении транспирации, с помощью которой из листьев водяными парами отводится излишек тепла, а также в создании оптимальных условий для вхождения углекислого газа через устьица. Многие хладнокровные животные (ящерицы, змеи, насекомые) используют солнечный свет для повышения температуры тела (некоторые змеи и ящерицы являются экологически теплокровными животными).
Одни факторы могут усиливать или ослаблять действие других факторов. Например, при оптимальной температуре повышается выносливость организмов к недостатку влаги и пищи; в свою очередь обилие пищи увеличивает устойчивость организмов к неблагоприятным климатическим обладать цветовым зрением, а у растений цветки имеют яркую окраску для привлечения насекомых-опылителей. Инфракрасные лучи с длиной волны более 0,750 мкм глаз человека не воспринимает, но они являются источником тепловой энергии (45% лучистой энергии). Эти лучи поглощаются тканями животных и растений, вследствие чего ткани нагреваются. У растений важнейшая функция инфракрасных лучей состоит в осуществлении транспирации, с помощью которой из листьев водяными парами отводится излишек тепла, а также в создании оптимальных условий для вхождения углекислого газа через устьица. Многие хладнокровные животные (ящерицы, змеи, насекомые) используют солнечный свет для повышения температуры тела (некоторые змеи и ящерицы являются экологически теплокровными животными).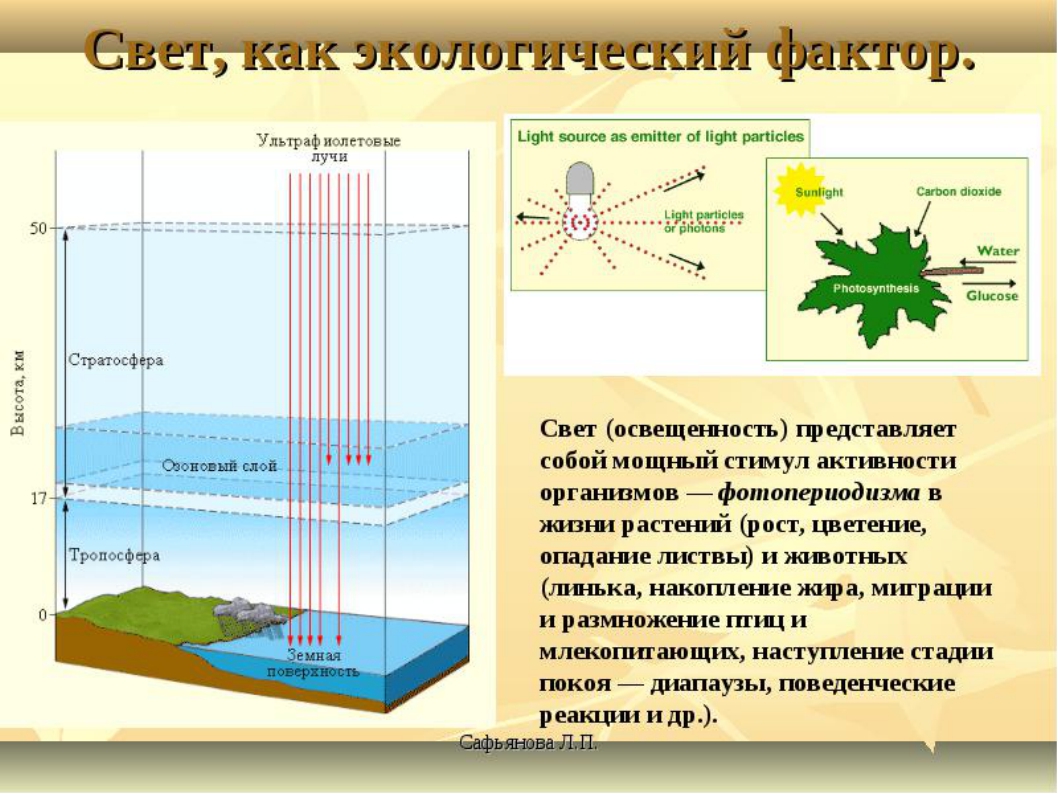 По отношению к свету различают несколько групп растений: 1. Светолюбивые — растения открытых пространств, на которые падает прямой свет. К ним относят растения степей, пустынь, полупустынь (ковыли, полыни, различные виды злаковых, например, пшеница и др.), а также растения верхних ярусов лесов (сосна, береза и др.). 2. Теневыносливые — растения, которые могут произрастать в условиях некоторого затенения (бук, дуб, граб, ель, липа, сирень и др.) Эта группа растений приспособлена к существованию в условиях недостаточного поступления света, так и в условиях хорошего освещения. Такие растения улавливают рассеянный свет темно-зелеными обогащенными хлорофиллом листьями. 3. Тенелюбивые — растения, которые не могут существовать в условиях попадания на них прямого света. К ним относятся растения, живущие под пологом леса: папоротники, звездчатка, ландыши и др. Приспособления организмов к световым факторам Растения Световые условия, связанные с вращением Земли, имеют отчетливую суточную и сезонную периодичность.
По отношению к свету различают несколько групп растений: 1. Светолюбивые — растения открытых пространств, на которые падает прямой свет. К ним относят растения степей, пустынь, полупустынь (ковыли, полыни, различные виды злаковых, например, пшеница и др.), а также растения верхних ярусов лесов (сосна, береза и др.). 2. Теневыносливые — растения, которые могут произрастать в условиях некоторого затенения (бук, дуб, граб, ель, липа, сирень и др.) Эта группа растений приспособлена к существованию в условиях недостаточного поступления света, так и в условиях хорошего освещения. Такие растения улавливают рассеянный свет темно-зелеными обогащенными хлорофиллом листьями. 3. Тенелюбивые — растения, которые не могут существовать в условиях попадания на них прямого света. К ним относятся растения, живущие под пологом леса: папоротники, звездчатка, ландыши и др. Приспособления организмов к световым факторам Растения Световые условия, связанные с вращением Земли, имеют отчетливую суточную и сезонную периодичность. Длина дня (фотопериод) имеет огромное значение в жизни растений и животных. Фотопериодизм — регуляция биоритма живых существ при помощи света. Различают суточный и сезонный фотопериодизм, а также периодизм процессов, протекающих на Солнце. Наиболее изучены суточный и сезонный фотопериодизм. “Биологические часы” определяют суточный ритм активности как целых организмов, так и процессов, происходящих даже на уровне клеток, в частности клеточных делений. У растений днем реализуются процессы световой фазы фотосинтеза и, частично, темновой фазы, а ночью — темновая фаза фотосинтеза. С фотопериодизмом у растений связано явление фототропизм — движение отдельных органов растения к свету, например, движение головки подсолнуха в течение дня по ходу движения Солнца, раскрытие соцветий одуванчика утром и закрытие их вечером, рост комнатных растений в освещенную сторону и т. д. (суточный фотопериодизм). У высших растений из-за укорочения светового дня и уменьшения интенсивности освещения происходит такое сезонное явление, как листопад.
Длина дня (фотопериод) имеет огромное значение в жизни растений и животных. Фотопериодизм — регуляция биоритма живых существ при помощи света. Различают суточный и сезонный фотопериодизм, а также периодизм процессов, протекающих на Солнце. Наиболее изучены суточный и сезонный фотопериодизм. “Биологические часы” определяют суточный ритм активности как целых организмов, так и процессов, происходящих даже на уровне клеток, в частности клеточных делений. У растений днем реализуются процессы световой фазы фотосинтеза и, частично, темновой фазы, а ночью — темновая фаза фотосинтеза. С фотопериодизмом у растений связано явление фототропизм — движение отдельных органов растения к свету, например, движение головки подсолнуха в течение дня по ходу движения Солнца, раскрытие соцветий одуванчика утром и закрытие их вечером, рост комнатных растений в освещенную сторону и т. д. (суточный фотопериодизм). У высших растений из-за укорочения светового дня и уменьшения интенсивности освещения происходит такое сезонное явление, как листопад. Свет оказывает влияние и на процесс развития растительных организмов. Некоторые растения эволюционно формировались при «коротком дне» (не более 12 часов в сутки), их называют растениями «короткого дня», а другие растения (они произрастают в средних и высоких широтах) — при «длинном дне» (продолжительность дня может достигать 20 часов и более), их называют растениями «длинного дня» (клюква, морошка и др.). Растения «длинного дня» не могут нормально развиваться на юге (они не дают семян), то же относится и к растениям «короткого дня», если их выращивать на севере, создавая все благоприятные условия, сохраняя продолжительность светлого времени суток. Животные Солнечная энергия непосредственно животными не усваивается, и, тем не менее, она является источником их жизнедеятельности. Кроме того, что солнечная энергия — источник жизни животных, она играет огромную роль в их жизни за счет следующих процессов. 1. Солнечный свет определяет суточный фотопериодизм жизни животных и их распределение по экологическим нишам. Различают животных, ведущих дневной и ночной образ жизни, что исключает конкуренцию за источники пищи. Большое значение свет играет и в жизни людей. Так, у некоторых людей наблюдается повышенная работоспособность в утренние часы («жаворонки»), а у других — в ночные часы («совы»). Солнечным днем эмоциональный настрой большинства людей значительно более высокий, чем в пасмурные или дождливые дни и т. д. 2. Солнечный свет позволяет животным легко ориентироваться в окружающей среде; свет эволюционно способствовал развитию органов зрения. абиотический фактор солнечный организм 3. Свет определяет и сезонный фотопериодизм, с которым связано изменение в ходе физиологических процессов (с наступлением осени интенсифицируется накопление запасных веществ в организме, меняется характер покровов и т. д.). Организмы, для которых характерны миграции (например, перелетные птицы), готовятся к ним и мигрируют, несмотря на наличие тепла и кормовой базы. Однако не все явления можно объяснить фотопериодизмом, например, миграции птиц из мест зимовки в теплые края, где длина дня сезонно не меняется, можно объяснить наличием «биологических часов», возникших в процессе эволюции и заложенных в генетическом коде. Обитатели подводных глубин находятся в постоянном мраке. Чем глубже проникает в море дневной свет, тем быстрее он слабеет. Различными приборами с фотографическими пластинками, которые намного чувствительней к свету, обнаружено, что свет проникает в толщу океана до глубины 1000 м. Глубже уже никакие приборы его не улавливают. Поэтому одни обитатели подводных глубин слепые, а у других глаза имеют особое строение, позволяющее улавливать даже малейшие проблески света. Их глаза достигают высокой степени развития. Иногда они бывают огромных размеров, снабженные системой линз. Другие животные имеют специальные приспособления, светящиеся различными цветами. Они не только освещают дорогу владельцу, но и приманивают добычу. А некоторые обитатели могут по своему желанию «тушить» и «зажигать» свет в этих органах. Также есть животные, (1 из видов кальмара), у которых в организме накапливается особая слизистая жидкость. Животное в момент опасности выпускает ее и скрывается от врага за светящимся голубым облаком.
Свет оказывает влияние и на процесс развития растительных организмов. Некоторые растения эволюционно формировались при «коротком дне» (не более 12 часов в сутки), их называют растениями «короткого дня», а другие растения (они произрастают в средних и высоких широтах) — при «длинном дне» (продолжительность дня может достигать 20 часов и более), их называют растениями «длинного дня» (клюква, морошка и др.). Растения «длинного дня» не могут нормально развиваться на юге (они не дают семян), то же относится и к растениям «короткого дня», если их выращивать на севере, создавая все благоприятные условия, сохраняя продолжительность светлого времени суток. Животные Солнечная энергия непосредственно животными не усваивается, и, тем не менее, она является источником их жизнедеятельности. Кроме того, что солнечная энергия — источник жизни животных, она играет огромную роль в их жизни за счет следующих процессов. 1. Солнечный свет определяет суточный фотопериодизм жизни животных и их распределение по экологическим нишам. Различают животных, ведущих дневной и ночной образ жизни, что исключает конкуренцию за источники пищи. Большое значение свет играет и в жизни людей. Так, у некоторых людей наблюдается повышенная работоспособность в утренние часы («жаворонки»), а у других — в ночные часы («совы»). Солнечным днем эмоциональный настрой большинства людей значительно более высокий, чем в пасмурные или дождливые дни и т. д. 2. Солнечный свет позволяет животным легко ориентироваться в окружающей среде; свет эволюционно способствовал развитию органов зрения. абиотический фактор солнечный организм 3. Свет определяет и сезонный фотопериодизм, с которым связано изменение в ходе физиологических процессов (с наступлением осени интенсифицируется накопление запасных веществ в организме, меняется характер покровов и т. д.). Организмы, для которых характерны миграции (например, перелетные птицы), готовятся к ним и мигрируют, несмотря на наличие тепла и кормовой базы. Однако не все явления можно объяснить фотопериодизмом, например, миграции птиц из мест зимовки в теплые края, где длина дня сезонно не меняется, можно объяснить наличием «биологических часов», возникших в процессе эволюции и заложенных в генетическом коде. Обитатели подводных глубин находятся в постоянном мраке. Чем глубже проникает в море дневной свет, тем быстрее он слабеет. Различными приборами с фотографическими пластинками, которые намного чувствительней к свету, обнаружено, что свет проникает в толщу океана до глубины 1000 м. Глубже уже никакие приборы его не улавливают. Поэтому одни обитатели подводных глубин слепые, а у других глаза имеют особое строение, позволяющее улавливать даже малейшие проблески света. Их глаза достигают высокой степени развития. Иногда они бывают огромных размеров, снабженные системой линз. Другие животные имеют специальные приспособления, светящиеся различными цветами. Они не только освещают дорогу владельцу, но и приманивают добычу. А некоторые обитатели могут по своему желанию «тушить» и «зажигать» свет в этих органах. Также есть животные, (1 из видов кальмара), у которых в организме накапливается особая слизистая жидкость. Животное в момент опасности выпускает ее и скрывается от врага за светящимся голубым облаком.
Архитектурное и музейное освещение — LightFactor
Архитектурное и музейное освещение — LightFactor | LightFactorПостоянная выставка
Ici a été fondée Montréal
Музей Пуэнт-а-Каллиер — Монреаль, 2017 Посмотреть проектАрхитектура
Monastère des Augustines
Квебек, 2015 Посмотреть проектВременная выставка
ИЛЛЮЗИИ, L’ART DE LA MAGIE
Музей Маккорда — Монреаль, 2017 Посмотреть проектВременная выставка
НАПОЛЕОН И ПАРИЖ
Канадский исторический музей — Оттава, 2015 Посмотреть проектАрхитектура
Au Sommet de la Place Ville-Marie
Ville-MariePlace — Монреаль, 2016 Посмотреть проектАрхитектура
АЛЬТ ОТЕЛЬ
Район Гриффинтаун — Монреаль, 2014 Посмотреть проект
Управляемый светом и кальцием фактор транскрипции для визуализации и манипулирования активированными нейронами
Tian, L.и другие. Визуализация нейронной активности у червей, мух и мышей с улучшенными кальциевыми показателями GCaMP. Nat. Методы 6 , 875–881 (2009).
CAS Статья Google ученый
Akerboom, J. et al. Оптимизация индикатора кальция GCaMP для визуализации нервной активности. J. Neurosci. 32 , 13819–13840 (2012).
CAS Статья Google ученый
Чен Т.-W. и другие. Сверхчувствительные флуоресцентные белки для визуализации активности нейронов. Природа 499 , 295–300 (2013).
CAS Статья Google ученый
Grynkiewicz, G., Poenie, M. & Tsien, R.Y. Новое поколение индикаторов Ca2 + со значительно улучшенными флуоресцентными свойствами. J. Biol. Chem. 260 , 3440–3450 (1985).
CAS PubMed Google ученый
Stosiek, C., Гаращук, О., Холтхофф, К. и Коннерт, А. Двухфотонная кальциевая визуализация нейронных сетей in vivo. Proc. Natl. Акад. Sci. США 100 , 7319–7324 (2003).
CAS Статья Google ученый
Fosque, B.F. et al. Нейронные цепи. Маркировка активных нейронных цепей in vivo с помощью разработанных кальциевых интеграторов. Наука 347 , 755–760 (2015).
CAS Статья Google ученый
Стрикленд, Д.и другие. Рациональное улучшение фотопереключателей на основе домена LOV. Nat. Методы 7 , 623–626 (2010).
CAS Статья Google ученый
Ву, Ю.И. и другие. Генетически кодируемый фотоактивируемый Rac контролирует подвижность живых клеток. Природа 461 , 104–108 (2009).
CAS Статья Google ученый
Барни, Г.и другие. Генетический дизайн сигнальных каскадов для регистрации активации рецепторов. Proc. Natl. Акад. Sci. США 105 , 64–69 (2008).
CAS Статья Google ученый
Inagaki, H.K. и другие. Визуализация нейромодуляции in vivo: TANGO-картирование передачи сигналов дофамина показывает, что аппетит контролирует чувствительность к сахару. Cell 148 , 583–595 (2012).
CAS Статья Google ученый
Капуст, р.Б., Тёзсер, Дж., Коупленд, Т.Д. и Во, Д.С. Специфичность P1 ‘протеазы вируса травления табака. Biochem. Биофиз. Res. Commun. 294 , 949–955 (2002).
CAS Статья Google ученый
Капуст, Р. Б. и др. Протеаза вируса травления табака: механизм автолиза и рациональный дизайн стабильных мутантов с каталитической способностью дикого типа. Protein Eng. 14 , 993–1000 (2001).
CAS Статья Google ученый
Hayashi-Takagi, A. et al. Маркировка и оптическое стирание следов синаптической памяти в моторной коре. Природа 525 , 333–338 (2015).
CAS Статья Google ученый
Пудасаини, А., Эль-Араб, К.К. & Zoltowski, B.D. Оптогенетические устройства на основе LOV: световые модули для передачи фоторегулируемого контроля клеточной передачи сигналов. Фронт. Мол. Biosci. 2 , 18 (2015).
Артикул Google ученый
Левская, А., Вайнер, О.Д., Лим, В.А., Фойгт, К.А. Пространственно-временной контроль клеточной передачи сигналов с использованием переключаемого светом белкового взаимодействия. Природа 461 , 997–1001 (2009).
CAS Статья Google ученый
Орт, П., Шнаппингер, Д., Hillen, W., Saenger, W. и Hinrichs, W. Структурные основы регуляции генов с помощью индуцируемой тетрациклином системы репрессор-оператор Tet. Nat. Struct. Биол. 7 , 215–219 (2000).
CAS Статья Google ученый
Park, H.Y. и другие. Конформационные изменения кальмодулина при связывании Са2 + изучали на микрофлюидном смесителе. Proc. Natl. Акад. Sci. США 105 , 542–547 (2008).
CAS Статья Google ученый
Konold, P.E. и другие. Разворачивание C-концевой спирали Jα в фоторецепторном домене LOV2, наблюдаемое с помощью колебательной спектроскопии с временным разрешением. J. Phys. Chem. Lett. 7 , 3472–3476 (2016).
CAS Статья Google ученый
Zayner, J.P. & Sosnick, T.R. Факторы, контролирующие химию фотоцикла домена LOV. PLoS One 9 , e87074 (2014).
Артикул Google ученый
Klapoetke, N.C. et al. Независимое оптическое возбуждение отдельных нейронных популяций. Nat. Методы 11 , 338–346 (2014).
CAS Статья Google ученый
Martell, J.D. et al. Разработал аскорбатпероксидазу как генетически кодируемый репортер для электронной микроскопии. Nat. Biotechnol. 30 , 1143–1148 (2012).
CAS Статья Google ученый
Лам, С.С. и др. Направленная эволюция APEX2 для электронной микроскопии и бесконтактной маркировки. Nat. Методы 12 , 51–54 (2015).
CAS Статья Google ученый
Rhee, H.W. и другие. Протеомное картирование митохондрий в живых клетках с помощью пространственно ограниченного ферментативного мечения. Наука 339 , 1328–1331 (2013).
CAS Статья Google ученый
Gradinaru, V. et al. Молекулярные и клеточные подходы для диверсификации и расширения оптогенетики. Cell 141 , 154–165 (2010).
CAS Статья Google ученый
Накашиба, Т., Янг, Дж. З., МакХью, Т. Дж., Буль, Д. Л.& Tonegawa, S. Трансгенное ингибирование синаптической передачи показывает роль выхода CA3 в обучении гиппокампа. Наука 319 , 1260–1264 (2008).
CAS Статья Google ученый
Армбрустер, Б.Н., Ли, X., Пауш, М.Х., Герлитце, С. и Рот, Б.Л. Доработка замка для соответствия ключу для создания семейства рецепторов, связанных с G-белком, активно активируемых инертным лигандом. Proc. Natl.Акад. Sci. США 104 , 5163–5168 (2007).
Артикул Google ученый
Wehr, M.C. и другие. Мониторинг регулируемых межбелковых взаимодействий с использованием расщепленного TEV. Nat. Методы 3 , 985–993 (2006).
CAS Статья Google ученый
Lee, D. et al. Временная маркировка и контроль нейромодуляторных цепей в головном мозге млекопитающих. Nat. Методы 14 , 495–503 (2017).
CAS Статья Google ученый
Лю, X. et al. Оптогенетическая стимуляция инграммы гиппокампа активирует воспоминание о страхе. Природа 484 , 381–385 (2012).
CAS Статья Google ученый
Guenthner, C.J., Miyamichi, K., Yang, H.H., Heller, H.К. и Луо, Л. Постоянный генетический доступ к временно активным нейронам через TRAP: целенаправленная рекомбинация в активных популяциях. Нейрон 78 , 773–784 (2013).
CAS Статья Google ученый
Реймерс Л.Г., Перкинс Б.Л., Мацуо Н. и Мэйфорд М. Локализация стабильного нейронного коррелята ассоциативной памяти. Наука 317 , 1230–1233 (2007).
CAS Статья Google ученый
Соренсен, А.T. et al. Надежная система маркировки активности для изучения активных нейронных ансамблей. eLife 5 , 2 (2016).
Артикул Google ученый
Файнберг, Э. и другие. GFP Reconstitution Across Synaptic Partners (GRASP) определяет клеточные контакты и синапсы в живых нервных системах. Нейрон 57 , 353–363 (2008).
CAS Статья Google ученый
Палмер, А.E. et al. Индикаторы Ca2 +, основанные на модернизированных с помощью вычислений парах кальмодулин-пептид. Chem. Биол. 13 , 521–530 (2006).
CAS Статья Google ученый
Халаваты, А.С. & Moffat, K. N- и C-концевые фланкирующие области модулируют индуцированную светом передачу сигнала в домене LOV2 фототропина 1 датчика синего света от Avena sativa. Биохимия 46 , 14001–14009 (2007).
CAS Статья Google ученый
Чао, Г. и др. Выделение и конструирование человеческих антител с использованием дрожжевой поверхности дисплея. Nat. Protoc. 1 , 755–768 (2006).
CAS Статья Google ученый
Зайнер, Дж. П., Антониу, К., Френч, А. Р., Хауз, Р. Дж. Младший и Сосник Т. Изучение моделей функции белков и аллостерии с широко распространенным мутационным анализом активируемого светом белка. Biophys. J. 105 , 1027–1036 (2013).
CAS Статья Google ученый
Zayner, J.P., Antoniou, C. & Sosnick, T.R. Аминоконцевая спираль модулирует активируемые светом конформационные изменения в AsLOV2. J. Mol. Биол. 419 , 61–74 (2012).
CAS Статья Google ученый
Конерманн, С.и другие. Оптический контроль эндогенной транскрипции и эпигенетических состояний млекопитающих. Природа 500 , 472–476 (2013).
CAS Статья Google ученый
Loh, K.H. и другие. Протеомный анализ неограниченных клеточных компартментов: синаптических щелей. Ячейка 166 , 1295–1307.e21 (2016).
CAS Статья Google ученый
Фактор дневного света
Факторы дневного света
Определение коэффициента дневного света
Коэффициент дневного света (DF) — очень распространенный и простой в использовании субъективное качество дневного света в комнате.Он описывает соотношение внешней освещенности над внутренней освещенностью, выраженной в процентах. Чем выше DF, тем больше естественного света в комнате. Выражается так:
DF = 100 * E дюйм / E доб
- E в
- внутренняя освещенность в фиксированной точке
- E доб
- За пределами горизонтальной освещенности при пасмурной погоде (небо CIE) или однородном небе.
E при освещенности можно рассматривать как сумму трех различных уровней освещенности:
- прямая освещенность, если небо видно из рассматриваемой точки (E D )
- освещенность за счет отражений от внешней среды (E ER )
- Освещенность за счет отражений на внутренних поверхностях (E IR )
Следовательно, коэффициент дневного света можно выразить как сумму трех составляющих:
DF = DC + ERC + IRC
- постоянного тока
- прямой компонент
- ERC
- компонент, отраженный извне
- IRC
- компонент с внутренним отражением
Коэффициент дневного света можно измерить для конкретной точки или выразить как среднее значение.Последний представляет собой среднее арифметическое суммы точечные измерения, сделанные на высоте 0,85 м в сетке, покрывающей всю площадь помещения. В разных странах действуют разные правила, и может потребоваться использование баллов. или средние измерения.
Коэффициент дневного света можно также выразить как среднее значение с помощью экспериментальных формул. Сегодня используется несколько формул для оценки средней плотности потока воздуха в комнате. В зависимости от страны и ее законодательства одно или другое может быть чаще встречается:
- Формула IES
- DF м, IES = (A окно ε U * 100) / (A этаж )
- Формула BRE
- DF м, BRE = (A окно α M t) / (A всего (1 — ρ м ²))
- Формула Сампнера
- DF м, Сампнер = (A окно α M t) / (2 A всего (1 — ρ м ))
- Законодательство Италии
- DF м, Италия = (A окно ε ψ t) / (A всего (1 — ρ м ))
- A окно
- Площадь окна без рамы, решеток и других препятствий [м²]
- A этаж
- Площадь помещения [м²]
- A всего
- Общая внутренняя площадь помещения [м²]
- ε
- Фактор для учета внешних препятствий
- U
- Коэффициент использования
- α
- Угол видимого неба из средней точки окна [°]
- M
- Коэффициент обслуживания окна
- т
- Коэффициент пропускания остекления
- ρ м
- Средний коэффициент отражения всех внутренних поверхностей
- ψ
- Коэффициент для учета толщины оконной стенки
Комнаты со средним DF 2% считаются освещенными дневным светом.Однако комната воспринимается как дневное освещение только тогда, когда DF выше 5%. Факторы дневного света всегда измеряются под пасмурным небом или в искусственное небо, имитирующее стандартное пасмурное небо CIE.
| Средний DF | Внешний вид | Энергетические последствия |
|---|---|---|
| комната выглядит мрачно | Электроосвещение необходимо большую часть дня | |
| от 2% до 5% | Внешний вид преимущественно дневного света, но дополнительно искусственный необходимо освещение. | Хороший баланс между освещением и тепловыми характеристиками |
| > 5% | Комната выглядит ярко освещенной | Дневное электрическое освещение требуется редко, но возможно тепловое проблемы из-за перегрева летом и тепловых потерь зимой |
В дополнение к формулам DF для комнат с боковым освещением, показанным выше, подробнее могут быть получены конкретные, такие как факторы дневного света для атриумных пространств.
Amazon.com: Светодиодная лампа Fotodiox Pro Factor Prizmo 300 RGB + W
Размер: 1×2
«Линия светильников FACTOR Prizmo сочетает в себе креативное освещение вещательного качества и доступность в мощном и портативном корпусе. Каждый светильник FACTOR Prizmo включает несколько встроенных режимов управления, включая режим CCT для белого света, регулируемый от 2800K до 10000K с регулируемыми точками пурпурного и зеленого , Режим HSI для доступа к полной цветовой гамме 360 ° и режим смешивания RBGW.Существует также предварительно запрограммированный режим фильтрации, который помогает подобрать стандартные гели, а также предварительно запрограммированные режимы специальных эффектов, включая эффекты полицейской машины, огня, вечеринки и фейерверков. Освещением FACTOR Prizmo можно управлять с помощью задней панели управления, контроллеров DMX512 или прилагаемого беспроводного пульта дистанционного управления 2,4 ГГц.
Фонари FACTOR Prizmo имеют прочную алюминиевую раму для долговечности и дополнительного теплоотвода. «Бесшумный» интеллектуальный вентилятор охлаждения гарантирует, что свет не перегреется, не создавая шума при съемке.Каждый фонарь FACTOR Prizmo поставляется в специально подобранном футляре на колесиках, что делает транспортировку безопасной и простой.
Светильник FACTOR Prizmo 1×2 потребляет максимум 300 Вт, поэтому вы можете использовать его в любой домашней электросети. Светильники FACTOR Prizmo также готовы к использованию в студии и на выезде благодаря возможности питания от батарей V-Lock (продаются отдельно). При использовании с сетевым питанием адаптер переменного тока предназначен для фиксации в пластине батареи с V-образным замком для надежного нескользящего соединения, удерживающего адаптер прикрепленным к световому блоку и не мешающим вам.Доступна дополнительная сотовая сетка 50 ° для направления и уменьшения разливов.
В комплект входят:
• FACTOR Prizmo 300 RGB + W Light Съемный дверной блок сарая
• Съемный софтбокс и решетка
• Монтажная скоба для 5/8 «» световой стойки
• Адаптер питания переменного тока 110-240 В и шнур
• Безопасность проводов Кабельные стяжки и кабельные стяжки
• Беспроводной пульт дистанционного управления 2,4 ГГц
• 3-контактный кабель дистанционного управления DMX
• Специальная сумка для переноски
• Руководство по эксплуатации »
Световое загрязнение как фактор рака груди и простаты
DOI: 10.1016 / j.scitotenv.2021.150918. Интернет впереди печати.Принадлежности Расширять
Принадлежности
- 1 Катедрас КОНАСИТ, 08400 Сьюдад-де-Мехико, Мексика; ICA, Словацкая академия наук, 845 03 Братислава, Словакия.Электронный адрес: [email protected].
- 2 МКА, Словацкая академия наук, 845 03 Братислава, Словакия; Факультет математики, физики и информатики, Университет Коменского, 842 48 Братислава, Словакия. Электронный адрес: [email protected].
- 3 UABC, Facultad de Ingeniería Arquitectura y Diseño, 22860 Энсенада, Мексика. Электронный адрес: [email protected].
- 4 Cátedras CONACYT, 08400 Ciudad de México, Мексика.Электронный адрес: [email protected].
- 5 Национальный информационный центр здравоохранения, 811 09 Братислава, Словакия. Электронный адрес: [email protected].
- 6 2-е отделение онкологии, Медицинский факультет Университета Коменского и Национальный институт рака, 833 10 Братислава, Словакия. Электронный адрес: [email protected].
- 7 Facultad de Ciencias Médicas, Национальный университет Куйо, 5500 Мендоса, Аргентина; Институт медицинской и экспериментальной биологии Куйо, UNCuyo CONICET, 5500 Мендоса, Аргентина.Электронный адрес: [email protected].
- 8 UABC, Facultad de Ingeniería Arquitectura y Diseño, 22860 Энсенада, Мексика. Электронный адрес: [email protected].
- 9 Facultad de Ciencias Médicas, Национальный университет Куйо, 5500 Мендоса, Аргентина; Институт медицинской и экспериментальной биологии Куйо, UNCuyo CONICET, 5500 Мендоса, Аргентина. Электронный адрес: [email protected].
Элемент в буфере обмена
Эктор Лэмпхар и др. Sci Total Environ. .
Показать детали Показать вариантыПоказать варианты
Формат АннотацияPubMedPMID
DOI: 10.1016 / j.scitotenv.2021.150918. Интернет впереди печати.Принадлежности
- 1 Катедрас КОНАСИТ, 08400 Сьюдад-де-Мехико, Мексика; ICA, Словацкая академия наук, 845 03 Братислава, Словакия. Электронный адрес: hasolanola @ conacyt.mx.
- 2 МКА, Словацкая академия наук, 845 03 Братислава, Словакия; Факультет математики, физики и информатики, Университет Коменского, 842 48 Братислава, Словакия. Электронный адрес: [email protected].
- 3 UABC, Facultad de Ingeniería Arquitectura y Diseño, 22860 Энсенада, Мексика. Электронный адрес: [email protected].
- 4 Cátedras CONACYT, 08400 Ciudad de México, Мексика.Электронный адрес: [email protected].
- 5 Национальный информационный центр здравоохранения, 811 09 Братислава, Словакия. Электронный адрес: [email protected].
- 6 2-е отделение онкологии, Медицинский факультет Университета Коменского и Национальный институт рака, 833 10 Братислава, Словакия. Электронный адрес: [email protected].
- 7 Facultad de Ciencias Médicas, Национальный университет Куйо, 5500 Мендоса, Аргентина; Институт медицинской и экспериментальной биологии Куйо, UNCuyo CONICET, 5500 Мендоса, Аргентина.Электронный адрес: [email protected].
- 8 UABC, Facultad de Ingeniería Arquitectura y Diseño, 22860 Энсенада, Мексика. Электронный адрес: [email protected].
- 9 Facultad de Ciencias Médicas, Национальный университет Куйо, 5500 Мендоса, Аргентина; Институт медицинской и экспериментальной биологии Куйо, UNCuyo CONICET, 5500 Мендоса, Аргентина. Электронный адрес: [email protected].
Элемент в буфере обмена
Полнотекстовые ссылки Опции дисплея CiteDisplayПоказать варианты
Формат АннотацияPubMedPMID
Абстрактный
Световое загрязнение — это глобальная экологическая проблема, затрагивающая светочувствительные организмы.Например, несколько исследователей признали подавление мелатонина у людей прямой причиной длительного воздействия высоких уровней искусственного освещения в ночное время. Другие определили низкий уровень мелатонина как фактор риска более высокой распространенности гормоночувствительного рака. В этой статье анализируется связь между световым загрязнением, оцененным по результатам анализа излучения спутниковых всемирных сборов ночного света с 1999 по 2012 год, и 25 025 случаев рака груди и 16 119 случаев рака простаты с 2003 по 2012 год.Оба типа рака увеличились в течение периода исследования, но световое загрязнение увеличилось в городских и пригородных районах и снизилось в сельской местности. Совокупное световое загрязнение за 5 лет показало положительную связь с раком груди, но не с раком простаты. Связь между световым загрязнением и раком груди сохранялась при приведении к стандартизованным по возрасту показателям со средним увеличением на 10,9 событий на 100 000 населения в год (95% доверительный интервал от 7,0 до 14,8). Мы пришли к выводу, что воздействие повышенного уровня светового загрязнения может быть фактором риска рака груди в Словакии.Эта работа может заинтересовать исследователей, изучающих взаимосвязь между атмосферными загрязнителями и растущей эпидемией рака. Результаты и методология могут быть экстраполированы на любую страну мира при наличии данных.
Ключевые слова: Рак молочной железы; Световое загрязнение; Подавление мелатонина; Рак простаты; Фактор риска.
Copyright © 2021 Elsevier B.V.Все права защищены.
Заявление о конфликте интересов
Заявление о конкурирующих интересах Авторы заявляют, что у них нет известных конкурирующих финансовых интересов или личных отношений, которые могли бы повлиять на работу, описанную в этой статье.
Похожие статьи
- Связь между искусственным освещением в ночное время и раком простаты в городе Кванджу и провинции Южная Чолла в Южной Корее.
Ким К.Й., Ли Э, Ким Й.Дж., Ким Дж. Ким KY и др. Chronobiol Int. 2017; 34 (2): 203-211. DOI: 10.1080 / 07420528.2016.1259241. Epub 2016 20 декабря. Chronobiol Int. 2017 г. PMID: 27996309
- Многогранное исследование загрязнения воздуха и смертности в Латинской Америке (исследование ESCALA).
Romieu I, Gouveia N, Cifuentes LA, de Leon AP, Junger W., Vera J, Strappa V, Hurtado-Díaz M, Miranda-Soberanis V, Rojas-Bracho L, Carbajal-Arroyo L, Tzintzun-Cervantes G; Комитет по надзору за здоровьем вуза.Romieu I, et al. Res Rep Health Eff Inst. 2012 Октябрь; (171): 5-86. Res Rep Health Eff Inst. 2012 г. PMID: 23311234
- Свет в большом городе: факторы риска рака груди у городских и сельских женщин в Израиле различаются.
Кешет-Ситтон А., Ор-Чен К., Ицхак С., Цабари И., Хаим А. Кешет-Ситтон А. и др. Integr Cancer Ther. 2017 июн; 16 (2): 176-187. DOI: 10.1177/1534735416660194. Epub 2016 20 июля. Integr Cancer Ther. 2017 г. PMID: 27440788 Бесплатная статья PMC.
- Суицидальные идеи.
Хармер Б., Ли С., Дуонг ТВХ, Саадабади А. Хармер Б. и др. 2021 6 августа. В: StatPearls [Интернет]. Остров сокровищ (Флорида): StatPearls Publishing; 2021 янв. 2021 6 августа. В: StatPearls [Интернет]. Остров сокровищ (Флорида): StatPearls Publishing; 2021 янв.PMID: 33351435 Бесплатные книги и документы. Рассмотрение.
- Искусственный свет ночью: мелатонин как посредник между окружающей средой и эпигеномом.
Хаим А., Зубидат А.Е. Хаим А. и др. Philos Trans R Soc Lond B Biol Sci. 2015 5 мая; 370 (1667): 20140121. DOI: 10.1098 / rstb.2014.0121. Philos Trans R Soc Lond B Biol Sci. 2015 г. PMID: 25780234 Бесплатная статья PMC.Рассмотрение.
Цитировать
КопироватьФормат: AMA APA ГНД NLM
СВЕТ КАК ЭКОЛОГИЧЕСКИЙ ФАКТОР ДЛЯ РАСТЕНИЙ
СВЕТ КАК ЭКОЛОГИЧЕСКИЙ ФАКТОР ДЛЯ РАСТЕНИЙ
Из всех климатических факторов, влияющих на развитие растений; CO2, температура, влажность и т. Д., свет — один из самых важных. Это важный фактор для растения, способный осуществлять фотосинтез, и от которого он напрямую зависит, чтобы синтезировать органическое вещество из минеральных веществ. Свет обеспечивает растение энергией, необходимой для фотосинтеза, который производит органическое вещество для его роста и развития.
Фотосинтетическая активность и ее эффективность возрастают с увеличением света, температуры и CO2, поэтому, если вы хотите добиться хорошего развития урожая и повышения урожайности, вы должны обеспечить хорошее и правильное освещение.Чем выше освещение, тем эффективнее фотосинтез и тем быстрее растение может развиваться. Однако с увеличением интенсивности света скорость фотосинтеза достигает предела. Эта точка максимума называется «точкой светового насыщения», при которой скорость физико-химического процесса фотосинтеза становится постоянной.
Количество света, получаемого растением, будет зависеть от двух основных факторов; интенсивность и количество часов ежедневного воздействия. Изменения в освещении, получаемом растениями, называются фотопериодом, и время воздействия или часы света, необходимые для развития растений и достижения цветения, варьируются в зависимости от вида.
Всем растениям нужен свет, и ниже определенного порога очень немногие из них способны выжить. Но и слишком много, и слишком мало света имеют пагубные последствия для растений. Растения, которые получают недостаточный уровень света, имеют меньший вегетативный рост, меньше цветут, становятся слабее и дают более мелкие листья. С другой стороны, растения, которые получают слишком много света, могут вызывать так называемый хлороз (хлороз — это пожелтение ткани листа, вызванное недостатком хлорофилла).
Этот жизненно важный параметр можно контролировать в теплице с помощью климатического программатора, который управляет затемняющими экранами (в случае высокой интенсивности света), затемнение (если нужно имитировать темноту, для короткодневных культур, когда они находятся при продолжительном дневном свете) и ассимиляционные огни (применяются, когда недостаточно света для хорошего роста сельскохозяйственных культур).
границ | Свет как фактор стресса для корней растений — случай корневого галотропизма
Свет как важный фактор окружающей среды для корней
В природе сидячие растения должны реагировать на суточные изменения световой среды. Одна из основных ролей света в жизни растений — обеспечивать энергией фотосинтез и регулировать развитие растений на различных стадиях, таких как прорастание семян, вегетативный рост, тропизмы и цветение. Известно, что фоторецепторы растений и родственные светочувствительные сигнальные молекулы участвуют в регуляции физиологических условий и морфологической пластичности в ответ на световую среду.Дарвин (1879) открыл отрицательный фототропизм корней растений. Год спустя Фрэнсис Дарвин и его отец Чарльз Дарвин опубликовали книгу «Сила движений растений». Они описали тропизмы как корней, так и побегов. Кроме того, они также предположили, что некоторая форма передачи сигналов на большие расстояния соединяет вершины сенсорных органов с активно растущими базальными частями (Darwin, 1879, 1880). С тех пор целенаправленная исследовательская работа в области физиологии растений открыла молекулу, передающую сигналы на большие расстояния, ауксин, что привело к пониманию фоторецепции растений.Такой ответ направленного роста на падающий свет называется фототропизмом. Положительный фототропизм, наблюдаемый в побегах, — это рост к источнику света; в то время как отрицательный фототропизм, наблюдаемый в корнях, отклоняется от источника света. Мы продемонстрировали, что короткое (10 с), но сильное (поток фотонов 82 мкмоль · м -2 с -1 ) синее свечение корней Arabidopsis вызывает немедленную генерацию активных форм кислорода (АФК) в область верхушки корня, что приводит к быстрому увеличению скорости роста корня (Yokawa et al., 2011, 2013). Этот активный ответ стимулированного светом роста корней называется тропизмом бегства (Xu et al., 2013; Yokawa et al., 2013; о корнях кукурузы см. Burbach et al., 2012). Такой тропизм позволит корням Arabidopsis избежать неблагоприятных световых условий, если выращивать на природе вне наших лабораторий.
Фоторецепторы в корнях
Было показано, что растение Arabidopsis экспрессирует 14 фоторецепторов, большинство из которых также присутствует в корнях (Briggs, Lin, 2012; Jeong, Choi, 2013; Briggs, 2014).Корни прорастают в темной почве, чтобы закрепить растение и впитать минеральные питательные вещества и воду. Сообщалось, что свет может проникать менее нескольких миллиметров из-за довольно высокого поглощения почвы (Woolley and Stoller, 1978). Тем не менее, небольшие трещины или механические удары часто могут происходить, что позволяет свету проникать глубже. Например, корни могут подвергаться воздействию света из-за резких перепадов температуры, землетрясения, сильного дождя, ветра и т. Д. Кроме того, очень важно, чтобы появляющийся корешок увеличивал скорость роста корня вскоре после прорастания семян на земле.Было необходимо развить способность корней реагировать на свет окружающей среды, когда в эволюции наземных растений появились первые цветковые растения с современной корневой системой. В следующем разделе мы обсудим интригующие взаимодействия между фитогормонами и связанными со светом сигнальными путями.
От актинового цитоскелета через рециклинг PIN2 к тропизму корней избегания соли
На клеточном уровне сообщалось, что белки PIN2 (PIN-FORMED 2; переносчик потока ауксина) в верхушках корней реагируют на световую среду (Laxmi et al., 2008). Ван и др. (2012) продемонстрировали, что базипетальный (направленный вверх) полярный транспорт ауксина на основе PIN2 подвергается контролю синего света, который регулирует отрицательный фототропизм корней Arabidopsis (Wan et al., 2012). Более того, Дьячок и др. (2011) сообщили, что активируемая светом COP1, убиквитинлигаза E3, способствует полимеризации актина и связыванию F-актина посредством регуляции нисходящего пути ARP2 / 3-SCAR в клетках корня. Это приводит к усилению роста корней в условиях освещения (Дьячок и др., 2011). Также сообщалось, что свет контролирует связывание F-актина в колеоптилях кукурузы (Waller and Nick, 1997), изменяя чувствительность клеток к ауксину, который отвечает за контроль F-актина, а также рост клеток (Nick et al., 2009 г.). Взаимодействие между транспортом F-актина и полярным ауксином, опосредованным рециклингом эндоцитарных пузырьков, особенно в переходной зоне верхушки корня, контролирует тропизм корня (Baluška et al., 1996, 2004, 2005, 2010; Baluška and Mancuso, 2013). Интересно, что предшественник эндогенного ауксина, индол-3-ацетальдегид (IAAld), вырабатывается неферментативно in vitro при освещении триптофаном в присутствии флавина, которого много в живых растительных клетках (Koshiba et al., 1993). Недавно мы предположили тесную связь между окислительно-восстановительным статусом и биосинтезом ауксина (ИУК) у растений (Yokawa et al., 2014). Взятые вместе, очевидно, что корни чрезвычайно чувствительны к воздействию света из-за присущей им эволюционной оптимизации для подземной жизни. Поэтому неудивительно, что освещенные корни молодых проростков Arabidopsis усиливают их рост с сопутствующим фототропизмом.
Несколько лет назад было показано, что корни Arabidopsis , подвергшиеся солевому стрессу, изменяют направление роста корней, чтобы избежать участков с высоким содержанием соли, благодаря так называемому тропизму избегания соли (Li and Zhang, 2008; Sun et al., 2008). Этот активный корневой тропизм требует пути восприятия ионного градиента, который затем будет контролировать изобилие, рециркуляцию и деградацию PIN2 (Li and Zhang, 2008; Sun et al., 2008). Это уникальное поведение корня Arabidopsis было связано с активностью фосфолипазы D Zeta2 (PLDζ2), которая стимулирует клатрин-опосредованный эндоцитоз PIN2, и этот тропизм также был назван корневым галотропизмом (Galvan-Ampudia et al., 2013; Rosquete and Kleine-Vehn, 2013; Пиерик, Тестеринк, 2014). Интересно, что аналогично, как и в случае галотропизма, PLDζ2 участвует в гидротропизме корней через опосредованное PIN2 подавление корневого гравитропизма (Taniguchi et al., 2010). Более того, PLDζ2 является критическим для рециклинга эндоцитов, чувствительного к brefeldin A, управляя рециклингом PIN2 (Li and Xue, 2007) и полярным транспортом ауксина в переходной зоне (Mancuso et al., 2007). Поскольку PIN2 имеет решающее значение в этом отношении, а также в адаптивных реакциях корней на свет (Laxmi et al., 2008; Sassi et al., 2012; Wan et al., 2012), очень важно проверить, подходят ли условия освещения. влияют на реакцию корней Arabidopsis на солевой стресс.
Растительные гормоны интегрированы со световыми сигнальными путями
Молекула Каррикина была выделена из дыма сгоревшего растительного сырья, и было обнаружено, что она сильно стимулирует прорастание семян растений (Flematti et al., 2004). Это соединение было идентифицировано как 3-метил-2H-фуро [2,3-c] пиран-2-он, каррикин-1 или каррикинолид-1 (KAR 1 ), показанные на рисунке 1A-a, и другие аналоги каррикинов. (KAR 2 -KAR 6 ) обычно обладают структурой бутенолида. Мутант Arabidopsis kai2 , лишенный генов KAI2 , нечувствительных к KAR 1 , запускал стимулирование прорастания семян. Считается, что KAI2 является предполагаемым рецептором каррикинов (Nelson et al., 2010; Waters et al., 2012). KAI2 является членом семейства α / β-гидролаз и также известен как ГИПОЧУВСТВИТЕЛЬНЫЙ К СВЕТУ (HTL). Роль KAI2 / HTL в световых ответах (рис. 1B) корней будет описана в следующем разделе.
РИСУНОК 1. Перекрестная связь между бутенолидными растительными гормонами и световыми сигнальными путями. (A) Химическая структура (а) Karrikin1, KAR 1 и (б) аналог стриголактона, GR24. (B) Взаимодействие гормональных и световых сигнальных путей.
Arabidopsis thaliana AtD14 (ортолог DWARF14) является паралогом KAI2 / HTL, который принадлежит к семейству α / β-гидролаз и играет роль в восприятии стриголактона (Waters et al., 2012). Стриголактоны синтезируются из каротиноидов и выделяются в корневые экссудаты для ускорения прорастания паразитических сорняков (Yoneyama et al., 2007), а также для стимуляции ветвления гиф у арбускулярных микоризных грибов (Akiyama et al., 2005). Стриголактоны действуют также как новый класс растительных гормонов, подавляющих рост пазушных почек (Umehara et al., 2008) и изменить корневую архитектуру (Колтай, 2011). Стриголактоны классифицируются как сесквитерпеновый лактон и имеют две части бутенолида (показаны на фиг.1A-b как синтезированный аналог GR24). Как описано выше, каррикины также содержат бутенолидные фрагменты. Следовательно, из-за структурного сходства двух молекул каррикинский рецептор KAI2 / HTL может отвечать на стриголактоны. Однако, напротив, предлагаемый рецептор стриголактона AtD14 нечувствителен к каррикинам; это указывает на то, что активность KAI2 / HTL как α / β-гидролазы более гибкая для определения структуры бутенолида, чем AtD14, вероятно, из-за эволюционных проблем (Waters et al., 2012).
Сообщалось, что белок F-бокса, БОЛЬШЕ AXILLARY BRANCHES2 (MAX2), расположен ниже по течению как KAI2 / HTL, так и AtD14 и, таким образом, необходим растениям для ответа на каррикин и стриголактон (Nelson et al., 2010; Waters et al., 2010; Waters et al. др., 2012). MAX2 образует SCF E3 убиквитин-лигазный комплекс (Skp1, Cullin1, F-box), который модулирует дальнейшие нижестоящие транскрипции (Stirnberg et al., 2007; Shen et al., 2012). Однако молекулярные функции регуляции морфогенеза растений с помощью KAI2 / HTL или AtD14 SCF MAX2 пока не известны.Интересно в этом отношении, что мутанты max2 сверхчувствительны к засухе и осмотическому стрессу, включая высокое содержание NaCl, маннита и глюкозы (Bu et al., 2014). Кроме того, стриголактоны играют положительную роль в адаптации растений к засухе и солевому стрессу (Ha et al., 2014). Мы обсудим эти новые аспекты в заключительном разделе.
Передача светового сигнала через COP1 и HY5 интегрирована с гормональной сигнализацией
В принципе, растения распознают свет как определенную длину волны с помощью определенных фоторецепторов.Активированные (возбужденные) фоторецепторы различными способами преобразуют световую информацию в физиологические сигналы. CONSTITUTIVE PHOTOMORPHOGENIC1 (COP1), убиквитинлигаза E3, участвует в световоспринимающем сигнальном каскаде, влияя на убиквитинирование целевых белков. Как только активность COP1 подавляется, нижестоящий фактор транскрипции bZIP, ELONGATED HYPOCOTYL5 (HY5) освобождается от убиквитинирования с помощью COP1 и запускает транскрипцию специфических генов, связанных со световыми ответами (Рисунок 1B слева).COP1 и HY5 контролируют рост корней, формирование боковых корней, рост кончиков корневых волосков, а также реакции прикосновения корней и гравитропизм (Oyama et al., 1997; Ang et al., 1998; Cluis et al., 2004; Datta et al. ., 2006, 2007; Сибут и др., 2006). Важно отметить, что SALTOLERANCE HOMOLOG2 (STh3) является взаимодействующим партнером COP1 и HY5, который контролирует корни и их уровни антоцианов (Datta et al., 2007).
Интересно, что воздействие стриголактона на растения подавляет активность COP1, что позволяет предположить, что стриголактоны могут имитировать восприятие света растениями (Tsuchiya et al., 2010). Они также продемонстрировали, что max2 мутант риса продуцирует избыток стриголактонов, что приводит к ингибированию COP1 и экспрессии светочувствительных генов (Tsuchiya et al., 2010). Кроме того, сообщалось, что мутант max2 сверхчувствителен к красному, дальнему красному и синему свету (Shen et al., 2007). О взаимодействии ауксина с сигнальным путем стриголактонов через MAX2 уже сообщалось (Hayward et al., 2009). Наконец, ключевому фактору транскрипции светового сигнала HY5 необходимы стриголактоны для стимуляции прорастания семян Arabidopsis во время термоингибитона (Toh et al., 2012). Свет также индуцирует биосинтез ауксина посредством фотовозбуждения флавинов (Koshiba et al., 1993; Yokawa et al., 2014).
Рецептор Каррикина KAI2 также известен как HTL. Экспрессия KAI2 / HTL сильно увеличивалась синим, красным и дальним красным светом, а HY5 связывается с промоторной областью HTL, указывая на то, что экспрессия KAI2 / HTL регулируется в ответ на световую среду (Sun and Ni, 2011). Более того, KAI2 / HTL располагается ниже HY5 (Рис. 1B) и индуцируется световой обработкой (Sun and Ni, 2011).Между тем, сообщалось, что обработка семян KAR 1 (химическая структура показана на рисунке 1A-a) улучшала световые реакции при прорастании и раннем развитии проростков (Nelson et al., 2010). Это означает, что Каррикины могут влиять на набор экспрессии генов в отношении световых ответов.
Корни растений, подвергшиеся стрессу засоления, увеличивают абсцизовую кислоту (ABA) и экспрессию генов, связанных с ABA (Raghavendra et al., 2010). АБК и стриголактоны синтезируются из каротиноидов, и уровни этих двух растительных гормонов влияют друг на друга (López-Ráez et al., 2010). Солевой стресс увеличивает уровень стриголактона в корнях, что затем способствует арбускулярному микоризному симбиозу, что приводит к изменению содержания АБК и снижению стрессовой реакции (Aroca et al., 2013). Стриголактоны, АБК и пути передачи сигналов цитокининов интегрированы, чтобы позволить растениям очень эффективно справляться с окружающей средой с высокой соленостью (Ha et al., 2014). Многие участники сигнальных путей фитогормона перекрываются и интегрируются с путями светового ответа.
Взятые вместе, эти находки предполагают, что гормональные и световые сигнальные пути используют одно и то же соединение для морфогенеза растений, а также для адаптации к абиотическим стрессам.Предлагаемые сигнальные пути (рис. 1B) сильно интегрированы, взаимодействуют друг с другом и, очевидно, представляют собой лишь «верхушку айсберга». В случае корней мы должны рассматривать свет исключительно как информацию (а не как источник энергии. ) в наше внимательное отношение к нашему пониманию фотоморфогенеза растений. На следующем занятии мы обсудим, как изменяется чувствительность корней к засолению при воздействии света на корни.
УФ-свет и УФР8 в ответах корней на засуху и засоление
Сообщалось, что снижение роста растений в условиях дефицита воды обусловлено фоторецептором УФ-В УФ-УСТОЙЧИВОСТЬ LOCUS 8 (UVR8; Kliebenstein et al., 2002; Браун и др., 2005; Favory et al., 2009; Fasano et al., 2014). Fasano et al. (2014) ранее показали, что ген UVR8 дополняет осмочувствительный мутант дрожжей mpk1 ppz1 . Экспрессия UVR8 была обнаружена в растениях Arabidopsis , выращенных в условиях солевого или осмотического стресса. Кроме того, эктопическая экспрессия UVR8 вызывает плейотропные эффекты на рост растений, такие как общее уменьшение размера органов растения, листья с более мелкими клетками, снижение роста корней и накопление флавоноидов.Это говорит о том, что морфогенные ответы УФ-В усиливаются у растений, гиперэкспрессирующих UVR8 и , выращенных при низких уровнях УФ-В-света. Дефекты роста растений, гиперэкспрессирующих UVR8 , еще более серьезны при осмотическом и солевом стрессе. Напротив, инактивация экспрессии UVR8 не влияет на рост побегов или корней в стандартных или умеренных условиях стресса засухи. Таким образом, гиперчувствительный ответ на осмотический стресс растений 35SUVR8 строго зависит от UVR8 (Fasano et al., 2014).
Имеются многочисленные свидетельства того, что осмотический стресс, а также воздействие ультрафиолетового излучения B более заметно на побегах, чем на корнях. При легком осмотическом стрессе корни продолжают расти (Bartels, Sunkar, 2005; Pardo, 2010; Werner et al., 2010). Более того, усиленный рост корней наблюдался у трансгенных растений с более высокой устойчивостью к засухе или солевому стрессу (Bartels and Sunkar, 2005). В то время как влияние осмотического стресса на развитие корней хорошо известно (недавно обзор Liu et al., 2014), влияние УФ-В на рост корней плохо изучено. В целом УФ-В снижает рост первичных корней проростков Arabidopsis (Kim et al., 1998; Tong et al., 2008). Сообщалось, что у взрослого растения повышенное выделение биомассы корнями происходит при стрессе УФ-В (Bussell et al., 2012).
Развитие более крупной корневой системы рассматривается как стратегия предотвращения засухи, которую растения применяют для улучшения поглощения воды и питательных веществ, когда их доступность в почве ограничена (Fukai and Cooper, 1995; Liao et al., 2001; Шарп и др., 2004; Пиерик, Тестеринк, 2014). Фоторецептор UV-B UVR8 экспрессируется в корнях растений дикого типа (Rizzini et al., 2011; обзор Yokawa and Baluška, 2014). Как показано на рисунке 2, UVR8 :: GFP экспрессируется в корнях Arabidopsis и транспортируется в ядра при облучении корней УФ-В. Избыточная экспрессия UVR8 значительно снижает рост корней при воздействии света и ухудшает их более слабые характеристики при осмотическом стрессе (Fasano et al., 2014). По сравнению с контрольными растениями плотность первичных и боковых корней составляла 13 и 60% соответственно.Это указывает на то, что рост боковых корней, зависимый от ауксина, был наиболее затруднен. Действительно, корневые фенотипы растений 35S-UVR8 напоминают фенотип ауксинового мутанта (Zhao, 2010). Анализ боковых корней показал, что количество зачатков боковых корней и появившихся боковых корней у растений 35S-UVR8 было на 12 и 68% соответственно меньше по сравнению с контрольными растениями (Fasano et al., 2014). Было обнаружено, что накопление флавоноидов в корне растений UVR8 со сверхэкспрессией увеличивалось в 2,2 раза, тогда как содержание конъюгатов ИУК имело тенденцию к снижению (Fasano et al., 2014). Таким образом, дефекты роста клеток корней 35S-UVR8 могут быть связаны с повышенными уровнями флавоноидов, которые, в свою очередь, изменяют полярный транспорт ауксина и / или гомеостаз ауксина (например, Santelia et al., 2008).
РИСУНОК 2. Перемещение UVR8 :: GFP в ядра в клетках корня Arabidopsis . (A) Область верхушки корня. (B) Увеличенное изображение клеток в переходной зоне. Полоса показывает 10 мкм. Arabidopsis корни подвергали обработке УФ-В в течение 1 часа (3.9 мкмоль м -2 с -1 ).
Флавоноиды синтезируются в корнях, регулируя ветвление корней, гравитропизм и адаптацию к стрессу (Brown et al., 2001; Buer, Muday, 2004; Agati et al., 2011; Emiliani et al., 2013). Концентрации флавоноидов значительно выше у растений, подвергшихся комбинированному стрессу, чем у растений, обработанных одним УФ-В (Hughes et al., 2010; Comont et al., 2012). Вместе все эти сообщения предполагают, что UVR8 , посредством контроля накопления флавоноидов, может быть обычным промежуточным звеном в путях передачи световых и гормональных сигналов для регулирования роста и развития корней в условиях абиотического стресса.Помимо УФ-B, засуха и засоление также связаны с накоплением антоцианов в различных тканях, включая корни (Agati et al., 2011; Emiliani et al., 2013; Meng, 2014). Важно в этом отношении, что индуцированная светом стимуляция роста корней Arabidopsis опосредуется посредством COP1-опосредованного накопления антоцианов (Meng, 2014).
Воздействие света на предотвращение засоления через галотропизм корня
Хотя известно, что верхушки корней, растущие в почве, находятся на переднем крае, подверженном солевому стрессу, влияние света на модуляцию чувствительности корней к засолению до сих пор не рассматривалось.Важно отметить, что в большинстве экспериментов с выращиванием в лаборатории сеянцев Arabidopsis используются корни, подвергающиеся воздействию света, хотя верхушки корней на природе в основном находятся в темноте.
Как показано на Рисунке 3, мы продемонстрировали, что воздействие света влияет на реакцию корня на воздействие засоления. Проростки Columbia WT выращивали на чашках с агаром, содержащих NaCl только в нижней части. С помощью этой системы можно наблюдать реакцию корневого тропизма на засоление, галотропизм.Наши методы следовали протоколу, описанному в (Galvan-Ampudia et al., 2013; Rosquete and Kleine-Vehn, 2013). После замены нижней части агара на NaCl-агар пластиковые чашки закрывали алюминиевой фольгой, чтобы обеспечить темную обработку. Период света / темноты составлял 16 ч / 8 ч, а интенсивность света составляла около 120 мкмоль м -2 с -1 , что сравнимо с условиями в Galvan-Ampudia et al. (2013). И темные, и светлые корни росли прямо к границе двух агаровых сред в контрольных экспериментах с 0 мМ NaCl (рис. 3).С другой стороны, корни, растущие в темноте, имеют более высокую чувствительность к NaCl, чем корни, подверженные воздействию света (таблица 1). Темно выращенные корни показали галотропизм, так как избегали участков с обогащенным солью агаром при всех концентрациях NaCl (рис. 3; табл. 1). При концентрации NaCl 100 мМ светлые корни продолжали врастать в соленую часть агара через границу между двумя частями агара даже через несколько дней. Это говорит о том, что свет либо мешает корням достичь галотропизма, либо что свет снижает чувствительность корней Arabidopsis к восприятию градиента солености.Тот факт, что выращенные на свету корни проявляли галотропизм при более высоких концентрациях NaCl (Таблица 1), предполагает, что свет мешает неуловимым корням ощущать градиенты Na + (Maathuis, 2014). Поскольку верхушки корней, растущих вне природы, обычно расположены глубоко в почве в темноте, можно ожидать, что их галотропизм более эффективен, чем для корней проростков, выращенных в лабораторных условиях.
РИСУНОК 3. Световая среда влияет на чувствительность корней Arabidopsis к засолению. Проростки Col-0 сначала выращивали на среде 1/2 MS в течение 5 дней. Нижнюю часть агара удаляли по диагонали и добавляли новую среду 1/2 MS, содержащую NaCl. Чашки Петри помещали в светлое или темное состояние на 3 дня. Корни, растущие на чашках с агаром в условиях темноты / света с 0, 100, 200 и 300 мМ NaCl в нижней половине чашек Петри. Метод, используемый для наложения корневого галотропизма, точно соответствует протоколу, опубликованному Galvan-Ampudia et al. (2013). Белые стрелки указывают вектор силы тяжести.Все сеянцы выращивали в вертикальном положении. Полоска показывает 1 см.
ТАБЛИЦА 1. Галотропные изгибы корней Arabidopsis , выращенных при различных условиях освещения.
Рост корней под действием света обходится дорого и меняет физиологию и морфологию целых проростков
Важно знать, что индуцированная светом стимуляция роста корней меняет физиологию целых проростков / растений. На рисунке 4 суммирован индуцированный светом рост корней, опосредованный взаимодействиями COP1 с актиновым цитоскелетом (Dyachok et al., 2011) и биосинтез антоцианов (Meng, 2014). И SCAR, и PAP-опосредованные пути требуют сахарозы в качестве источника энергии (Kurata and Yamamoto, 1997; Kircher and Schopfer, 2012; Maier et al., 2013; Meng, 2014). Корни, лишенные коактиватора транскрипции ANGUSTIFOLIA3 (AN3), имеют более низкие уровни антоцианов и более длинные корни на свету, чем корни WT (Meng, 2014). Напротив, мутантные корни an3 короче корней WT при выращивании в темноте. Интересно, что AN3 связывается с промотором COP1, чтобы ингибировать индуцированное светом удлинение корня (Meng, 2014).Помимо антоцианов, облученные светом корни также демонстрируют довольно резкое усиление метаболизма фенилпропаноидов, вызывая не только флавоноиды, но и монолигнолглюкозиды (Hemm et al., 2004). Возможное влияние света на формирование полос Каспариана в энтодерме можно ожидать и легко проверить.
Индуцированная светом полимеризация актина, полярный транспорт ауксина, рост корней и биосинтез антоцианов — это энергоемкие процессы. Повышенная потребность в сахарозе для фотоморфогенеза корней (Costigan et al., 2011; Варнасоория и Монтгомери, 2011; Yokawa et al., 2013) происходит за счет других потенциальных стоков тела растения (Kurata, Yamamoto, 1997; Yazdanbakhsh et al., 2011; Kircher, Schopfer, 2012). Более того, солевой стресс, воспринимаемый локально на верхушке корня, быстро распространяется по телу растения посредством системной передачи сигналов (Choi et al., 2014; Gilroy et al., 2014). Поэтому важно знать, что освещение корней имеет довольно драматические последствия для физиологии и морфологии всего саженца.
Выводы и перспективы
Наши результаты с использованием солевого стресса показывают, что корни, подвергшиеся воздействию света, демонстрируют различное поведение и реакцию при солевом стрессе. Один из возможных сценариев, основанный на сообщениях о том, что освещенные корни повышают уровень стриголактона (Koltai et al., 2011), а стриголактон увеличивает засухоустойчивость и солеустойчивость корней (Ha et al., 2014), заключается в том, что освещенные корни менее чувствительны к солевой стресс. В то время как темные корни проявляют галотропизм (тропизм избегания соли) за счет роста вокруг частей агара, обогащенного солью, освещенные светом корни вырастают в агар, обогащенный солью, если уровни NaCl не слишком высоки (таблицы 1 и 2).Поскольку индуцированные светом уровни стриголактона изменяют также ответы корней на низкий уровень фосфатов (Mayzlish-Gati et al., 2012), можно предположить, что воздействие света на корни Arabidopsis изменяет всю их физиологию. Фактически, наши предварительные данные показывают, что корни Arabidopsis , подвергшиеся воздействию света, также изменяют чувствительность к токсичности алюминия (Kagenishi et al., В стадии подготовки). Кроме того, уровни ИУК в освещенных (1 ч) корнях кукурузы были более чем вдвое выше по сравнению с контрольными уровнями (Yokawa et al., в процессе подготовки). Интересно, что повышенный уровень стриголактона напоминает воздействие света на корни. Было показано, что воздействие света на актиновый цитоскелет, а также увеличение количества и рециклинга переносчика ауксина PIN2 (Laxmi et al., 2008; Dyachok et al., 2011; Wan et al., 2012; Pandya-Kumar et al., 2014) ). Кроме того, солевой стресс, стресс от засухи, холодовой стресс, щелочной стресс, токсичность алюминия; все эти проблемы нацелены на транспортер ауксина PIN2, который экспрессируется в переходной зоне верхушки корня (обсуждается у Baluška et al., 2010; Балушка и Манкузо, 2013). Во избежание засветки корней мы рекомендуем использовать частично затемненные чашки Петри (Yokawa et al., 2011, 2013; Xu et al., 2013). Это важно, поскольку воздействие на корни светом изменяет не только реакцию корней на различные стрессы, но и общую физиологию таких сеянцев. Например, сообщалось, что циркадные ритмы в корнях Arabidopsis регулируются частью побега (James et al., 2008). Важно отметить, что передача сигналов и гомеостаз АФК является основным модулятором циркадных ритмов как у прокариот, так и у эукариот (Edgar et al., 2012; Lai et al., 2012; Стангерлин и Редди, 2013). Возможно, что сеянцы изменят даже свои циркадные часы, когда корни будут освещены.
ТАБЛИЦА 2. Arabidopsis длины корней в различных условиях соли и освещения.
Хотя корни растений являются гетеротрофными органами растений, они требуют активного подавления экспрессии фотосинтетических генов. Белок тирозилпротеинсульфотрансферазы (TPST) HPS7 недавно стал ключевым игроком, контролирующим это активное подавление (Kang et al., 2014). Интересно, что hsp7 мутантных корней демонстрируют усиленные эффекты, связанные с фотосинтезом, в условиях стресса, вызванного дефицитом фосфата. Предыдущие исследования идентифицировали золотоподобные транскрипционные факторы GLK1 и GLK2, участвующие в активации фотосинтетических генов в корнях (Waters et al., 2009; Kobayashi et al., 2013). Как hsp7, , так и мутантные корни GLK-ox не обнаруживают задержки роста корней и каких-либо других фенотипов при выращивании в темноте (Kang et al., 2014).
Наконец, с точки зрения физиологии растений индуцированный светом рост корней у Arabidopsis рассматривает высокую скорость роста корней как признак оптимальных условий для роста корней.Напротив, нейробиологическая интерпретация растительной нейробиологии этой повышенной скорости роста корней заключается в том, что подвергшиеся воздействию света корни испытывают стресс и что такие стрессированные корни пытаются избежать этой неблагоприятной ситуации (Yokawa et al., 2011, 2013; Xu et al. ., 2013). Alpi et al. (2007) критики инициативы нейробиологии растений утверждали, что «… нейробиология растений не способствует нашему пониманию физиологии растений, биологии растительных клеток или передачи сигналов». Теперь, примерно 7 лет спустя, становится очевидным, что растения и их корни являются поведенчески намного сложнее, чем это предполагалось в рамках классической физиологии растений (Trewavas, 2005, 2009, 2014; Trewavas, Baluška, 2011; Baluška, Mancuso, 2013; Pierik and Testerink, 2014).Освещение корней является обычной лабораторной практикой с тех пор, как Arabidopsis thaliana был введен в науку о растениях в качестве модельного организма. Однако важно знать, что воздействие света на корни становится стрессовым фактором для выращенных в лаборатории сеянцев Arabidopsis . Это наглядный пример полезности рассмотрения растений как активно живущих и чувствительных организмов, разумно решающих свои специфические для растений проблемы.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Кен Йокава был поддержан докторской стипендией JSPS (Японское общество содействия науке). Мы благодарим профессора Гарета И. Дженкинса (Университет Глазго) за предоставленные нам семена линии UVR8 :: GFP.
Список литературы
Агати, Г., Бириколти, С., Гуиди, Л., Феррини, Ф., Фини, А., и Таттини, М. (2011). Биосинтез флавоноидов аналогичным образом усиливается УФ-излучением и засолением корневой зоны у листьев L. vulgare.J. Plant Physiol. 168, 204–212. DOI: 10.1016 / j.jplph.2010.07.016
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Alpi, A., Amrhein, N., Bertl, A., Blatt, M. R., Blumwald, E., Cervone, F., et al. (2007). Нейробиология растений: нет мозга — нет выгоды? Trends Plant Sci. 12, 135–136. DOI: 10.1016 / j.tplants.2007.03.002
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Ang, L.H., Chattopadhyay, S., Wei, N., Oyama, T., Okada, K., Batschauer, A., et al. (1998). Молекулярное взаимодействие между COP1 и HY5 определяет регуляторный переключатель для управления светом развития Arabidopsis . Мол. Cell 1, 213–222. DOI: 10.1016 / S1097-2765 (00) 80022-2
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Арока, Р., Руис-Лозано, Дж. М., Замаррено, А. М., Паз, Дж. А., Гарсия-Мина, Дж. М., Посо, М. Дж. И др. (2013). Симбиоз арбускулярной микоризы влияет на выработку стриголактона при засолении и снимает солевой стресс у растений салата. J. Plant Physiol. 170, 47–55. DOI: 10.1016 / j.jplph.2012.08.020
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Бай, С., Яо, Т., Ли, М., Го, X., Чжан, Ю., Чжу, С. и др. (2014). PIF3 участвует в подавлении роста первичных корней Arabidopsis , индуцированном оксидом азота на свету. Мол. Завод 7, 616–625. DOI: 10.1093 / mp / sst142
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Балушка, Ф., Манкузо, С., Фолькманн, Д., и Барлоу, П. В. (2004). Верхушки корня как командные центры растений: уникальный «мозговой» статус переходной зоны верхушки корня. Biologia 59, 9–17.
Google Scholar
Бартельс, Д., Сункар, Р. (2005). Засухо- и солеустойчивость растений. Crit. Rev. Plant Sci. 24, 23–58. DOI: 10.1080 / 073526805410
CrossRef Полный текст | Google Scholar
Браун, Б. А., Клуа, К., Цзян, Г. Х., Кайзерли, Э., Herzyk, P., Kliebenstein, D. J., et al. (2005). Сигнальный компонент, специфичный для УФ-В, управляет защитой растений от УФ-излучения. Proc. Natl. Акад. Sci. США 102, 18225–18230. DOI: 10.1073 / pnas.0507187102
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Браун Д. Э., Рашотт А. М., Мерфи А. С., Норманли Дж., Тейг Б. В., Пир В. А. и др. (2001). Флавоноиды действуют как негативные регуляторы транспорта ауксина in vivo у Arabidopsis . Plant Physiol. 126, 524–535. DOI: 10.1104 / стр. 126.2.524
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Bu, Q., Lv, T., Shen, H., Luong, P., Wang, J., Wang, Z., et al. (2014). Регулирование засухоустойчивости с помощью F-box белка MAX2 в Arabidopsis . Plant Physiol. 164, 424–439. DOI: 10.1104 / стр.113.226837
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Бурбах, К., Маркус, К., Чжан, Ю., Шлихт, М., и Балушка, Ф. (2012). Светобоязнь корней кукурузы. Завод Сигнал. Behav. 7, 874–878. DOI: 10.4161 / psb.21012
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Басселл, Дж. С., Гвинн-Джонс, Д., Гриффит, Г. У., и Скаллион, Дж. (2012). Ответы Calamagrostis purpurea над землей и под землей на УФ-В излучение и повышенный уровень CO 2 при ограничении фосфора. Physiol.Растение. 145, 619–628. DOI: 10.1111 / j.1399-3054.2012.01595.x
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Чой, В. Г., Тойота, М., Ким, С. Х., Хиллари, Р., и Гилрой, С. (2014). Вызванные солевым стрессом волны Ca 2+ связаны с быстрой передачей сигналов от корня к побегам на большие расстояния. Proc. Natl. Акад. Sci. США 111, 6497–6502. DOI: 10.1073 / pnas.1319955111
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Комонт, Д., Уинтерс, А., Гвинн-Джонс, Д. (2012). Акклимация и взаимодействие между засухой и повышенным УФ-B у A. thaliana : различия в реакции на лечение, восстановление и размножение. Ecol. Evol. 2, 2695–2709. DOI: 10.1002 / ece3.387
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Костиган, С. Е., Варнасоория, С. Н., Хамфрис, Б. А., и Монтгомери, Б. Л. (2011). Локализованный в корне синтез фитохромного хромофора необходим для фоторегуляции удлинения корня и влияет на чувствительность корней к жасмоновой кислоте у Arabidopsis . Plant Physiol. 157, 1138–1150. DOI: 10.1104 / стр.111.184689
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Дарвин, К. Р. (1880). Сила движения растений . Лондон: Джон Мюррей.
Google Scholar
Дарвин, Ф. (1879). Über das negativ wachstum heliotropischer wurzeln im licht und im finstern. Арб. Д. Бот. Inst. Würzburg 2, 521–528.
Датта, С., Хеттиараччи, К., Йоханссон, Х., и Холм, М. (2007). СОЛЕНОСТОЙЧИВОСТЬ HOMOLOG2, белок B-бокса в Arabidopsis , который активирует транскрипцию и положительно регулирует опосредованное светом развитие. Растительная клетка 19, 3242–3255. DOI: 10.1105 / tpc.107.054791
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Дьячок, Дж., Чжу, Л., Ляо, Ф., Хе, Дж., Хук, Э. и Бланкафлор, Э. Б. (2011). SCAR опосредует индуцированное светом удлинение корня у Arabidopsis через фоторецепторы и протеасомы. Растительная клетка 23, 3610–3626. DOI: 10.1105 / tpc.111.088823
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Эдгар, Р.С., Грин, Э.В., Чжао, Ю., ван Оойен, Г., Ольмедо, М., Цинь, X. и др. (2012). Пероксиредоксины — консервативные маркеры циркадных ритмов. Природа 485, 459–464. DOI: 10.1038 / природа11088
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Фазано Р., Гонсалес Н., Tosco, A., Piaz, F. D., Docimo, T., Serrano, R., et al. (2014). Роль Arabidopsi s – UV RESISTANCE LOCUS 8 в замедлении роста растений при осмотическом стрессе и низких уровнях УФ-B. Мол. Завод 5, 773–791. DOI: 10.1093 / mp / ssu002
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Favory, J. J., Stec, A., Gruber, H., Rizzini, L., Oravecz, A., Funk, M., et al. (2009). Взаимодействие COP1 и UVR8 регулирует UV-B-индуцированный фотоморфогенез и акклиматизацию к стрессу у Arabidopsis . EMBO J. 28, 591–601. DOI: 10.1038 / emboj.2009.4
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Фукаи, С., Купер, М. (1995). Создание засухоустойчивых сортов риса по физиоморфологическим признакам. Field Crops Res. 40, 67–86. DOI: 10.1016 / 0378-4290 (94) 00096-U
CrossRef Полный текст | Google Scholar
Гальван-Ампудиа, К.С., Юлковска, М.М., Дарвиш, Э., Гандулло, Дж., Корвер, Р.A., Brunoud, G., et al. (2013). Галотропизм — это реакция корней растений на избегание засоления. Curr. Биол. 23, 2044–2050. DOI: 10.1016 / j.cub.2013.08.042
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Гилрой, С., Сузуки, Н., Миллер, Г., Чой, В. Г., Тойота, М., Девиредди, А. Р. и др. (2014). Приливная волна сигналов: кальций и АФК на переднем крае быстрой системной передачи сигналов. Trends Plant Sci. 19, 623–630.DOI: 10.1016 / j.tplants.2014.06.013
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Го Ю., Чжэн З., Ла Клер Дж. Дж., Чори Дж. И Ноэль Дж. П. (2013). Восприятие каррикина из дыма α / β-гидролазой KAI2 из Arabidopsis . Proc. Natl. Акад. Sci. США 110, 8284–8289. DOI: 10.1073 / pnas.1306265110
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
га, C.В., Лейва-Гонсалес, М. А., Осакабе, Ю., Тран, У. Т., Нишияма, Р., Ватанабе, Ю., и др. (2014). Положительная регулирующая роль стриголактона в ответах растений на засуху и солевой стресс. Proc. Natl. Акад. Sci. США 111, 851–856. DOI: 10.1073 / pnas.1322135111
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Хемм, М. Р., Райдер, С. Д., Огас, Дж., Мерри, Д. Дж. И Чаппл, К. (2004). Свет вызывает метаболизм фенилпропаноидов в корнях Arabidopsis . Плант Дж. 38, 765–778. DOI: 10.1111 / j.1365-313X.2004.02089.x
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Хьюз, Н. М., Рейнхард, К., Фейлд, Т. С., Герарди, А. Р., и Смит, В. К. (2010). Связь между производством антоцианов зимой и стрессом от засухи у вечнозеленых видов покрытосеменных. J. Exp. Бот. 61, 1699–1709. DOI: 10.1093 / jxb / erq042
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Джеймс, А.Б., Монреаль, Дж. А., Ниммо, Г. А., Келли, К. Л., Херзик, П., Дженкинс, Г. И. и др. (2008). Циркадные часы в корнях Arabidopsis — это упрощенная подчиненная версия часов в побегах. Наука 322, 1832–1835. DOI: 10.1126 / science.1161403
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Кан, Дж., Ю, Х., Тиан, К., Чжоу, В., Ли, К., Цзяо, Ю. и др. (2014). Подавление экспрессии фотосинтетических генов в корнях необходимо для устойчивого роста корней при дефиците фосфатов. Plant Physiol. 165, 1156–1170. DOI: 10.1104 / стр.114.238725
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Кирхер, С., Шопфер, П. (2012). Фотосинтетическая сахароза действует как происходящий из семядолей сигнал на большие расстояния, чтобы контролировать рост корней во время раннего развития проростков у Arabidopsis . Proc. Natl. Акад. Sci. США 109, 11217–11221. DOI: 10.1073 / pnas.1203746109
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Клебенштейн, Д.Дж., Лим, Дж. Э., Ландри, Л. Г., и Ласт, Р. Л. (2002). Arabidopsis UVR8 регулирует передачу и толерантность ультрафиолетового B-сигнала и содержит последовательность, сходную с человеческим регулятором конденсации хроматина. Plant Physiol. 130, 234–243. DOI: 10.1104 / pp.005041
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Кобаяси, К., Сасаки, Д., Ногучи, К., Фудзинума, Д., Комацу, Х., Кобаяши, М., и др. (2013). Фотосинтез хлоропластов корня развился в линиях Arabidopsis , сверхэкспрессирующих факторы транскрипции GOLDEN2-LIKE. Physiol растительных клеток. 54, 1365–1377. DOI: 10.1093 / pcp / pct086
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Колтай, Х. (2011). Стриголактоны — регуляторы развития корней. New Phytol. 190, 545–549. DOI: 10.1111 / j.1469-8137.2011.03678.x
CrossRef Полный текст | Google Scholar
Koltai, H., Cohen, M., Chesin, O., Mayzlish-Gati, E., Bécard, G., Puech, V., et al. (2011). Свет является положительным регулятором уровня стриголактона в корнях томатов. J. Plant Physiol. 168, 1993–1996. DOI: 10.1016 / j.jplph.2011.05.022
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Кошиба Т., Ямаути К., Мацуяма Х., Миякадо М., Сори И. и Сато М. (1993). Флавин-фотосенсибилизированное производство индол-3-ацетальдегида из триптофана. Tetrahedron Lett. 34, 7603–7606. DOI: 10.1016 / S0040-4039 (00) 60411-2
CrossRef Полный текст | Google Scholar
Курата, Т., и Ямамото, К. Т. (1997). Растяжение корней, стимулированное светом, у Arabidopsis thaliana . J. Plant Physiol. 151, 346–351. DOI: 10.1016 / S0176-1617 (97) 80263-5
CrossRef Полный текст | Google Scholar
Лай, А.Г., Доэрти, К.Дж., Мюллер-Робер, Б., Кей, С.А., Шипперс, Дж. Х., и Дейквел, П. П. (2012). CIRCADIAN CLOCK-ASSOCIATED 1 регулирует гомеостаз АФК и реакции на окислительный стресс. Proc. Natl. Акад. Sci. США 109, 17129–17134.DOI: 10.1073 / pnas.12009
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Ляо, Х., Рубио, Г., Янь, X., Цао, А., Браун, К. М., и Линч, Дж. П. (2001). Влияние доступности фосфора на глубину прикорневой части бобов обыкновенной. Почва растений 232, 69–79. DOI: 10.1023 / A: 1010381919003
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Лопес-Раэс, Дж. А., Колен, В., Чарнихова, Т., Малдер, П., Ундас, А. К., сержант, М. Дж. И др. (2010). Влияет ли абсцизовая кислота на биосинтез стриголактона? New Phytol. 187, 343–354. DOI: 10.1111 / j.1469-8137.2010.03291.x
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Maier, A., Schrader, A., Kokkelink, L., Falke, C., Welter, B., Iniesto, E., et al. (2013). Light и убиквитинлигаза E3 COP1 / SPA контролируют стабильность белков факторов транскрипции MYB PAP1 и PAP2, участвующих в накоплении антоцианов в Arabidopsis . Plant J. 74, 638–651. DOI: 10.1111 / tpj.12153
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Манкузо, С., Маррас, А. М., Мугнаи, С., Шлихт, М., Зарски, В., Ли, Г. и др. (2007). Фосфолипаза dzeta2 управляет везикулярной секрецией ауксина для его полярного межклеточного транспорта в переходной зоне верхушки корня. Завод Сигнал. Behav. 2, 240–244. DOI: 10.4161 / psb.2.4.4566
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Майзлиш-Гати, Э., De-Cuyper, C., Goormachtig, S., Beeckman, T., Vuylsteke, M., Brewer, P. B., et al. (2012). Стриголактоны участвуют в ответной реакции корней на низкие фосфатные условия у Arabidopsis . Plant Physiol. 160, 1329–1341. DOI: 10.1104 / стр.112.202358
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Менг, Л. С. (2014). Коактиватор транскрипции Arabidopsis ANGUSTIFOLIA3 модулирует накопление антоцианов и индуцированное светом удлинение корня посредством трансрепрессии конститутивного фотоморфогенного 1. Среда растительных клеток. doi: 10.1111 / pce.12456 [Epub перед печатью].
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Нельсон, Д. К., Флематти, Г. Р., Райзборо, Дж. А., Гизалберти, Э. Л., Диксон, К. В., и Смит, С. М. (2010). Каррикинс усиливает световые реакции во время прорастания и развития проростков у Arabidopsis thaliana . Proc. Natl. Акад. Sci. США 107, 7095–7100. DOI: 10.1073 / pnas.05107
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Пандья-Кумар, Н., Шема, Р., Кумар, М., Майзлиш-Гати, Э., Леви, Д., Земах, Х. и др. (2014). Аналог стриголактона GR24 запускает изменения полярности PIN2, транспорта пузырьков и архитектуры актиновых филаментов. New Phytol. 202, 1184–1196. DOI: 10.1111 / Nph.12744
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Rizzini, L., Favory, J. J., Cloix, C., Faggionato, D., O’Hara, A., Kaiserli, E., et al. (2011). Восприятие УФ-В белком Arabidopsis UVR8. Наука 332, 103–106. DOI: 10.1126 / science.1200660
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Сантелия, Д., Хенрикс, С., Винченцетти, В., Зауэр, М., Биглер, Л., Кляйн, М. и др. (2008). Флавоноиды перенаправляют опосредованные PIN-кодами полярные потоки ауксина во время корневых гравитропных реакций. J. Biol. Chem. 283, 31218–31226. DOI: 10.1074 / jbc.M710122200
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Сасси, М., Лу, Ю., Чжан, Ю., Ван, Дж., Донукше, П., Блилоу, И. и др. (2012). COP1 обеспечивает координацию роста корней и побегов с помощью света посредством модуляции PIN1 и PIN2-зависимого транспорта ауксина у Arabidopsis . Разработка 139, 3402–3412. DOI: 10.1242 / dev.078212
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Шарп Р. Э., Поройко В., Хейлек Л. Г., Споллен В. Г., Спрингер Г. К., Бонерт, Х. Дж. И др. (2004).Поддержание роста корней при дефиците воды: от физиологии до функциональной геномики. J. Exp. Бот. 55, 2343–2351. DOI: 10.1093 / jxb / erh376
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Сибут, Р., Сукумар, П., Хеттиараччи, К., Холм, М., Мудай, Г. К., и Хардтке, К. С. (2006). Противоположные фенотипы роста корней у мутантов hy5 по сравнению с hy5 hyh коррелируют с усилением конститутивной передачи сигналов ауксина. PLoS Genet. 2: e202. DOI: 10.1371 / journal.pgen.0020202
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Sun, F., Zhang, W., Hu, H., Li, B., Wang, Y., Zhao, Y., et al. (2008). Соль модулирует путь передачи сигналов силы тяжести для регулирования направления роста первичных корней у Arabidopsis . Plant Physiol. 146, 178–188. DOI: 10.1104 / стр.107.109413
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Танигучи, Ю.Ю., Танигучи, М., Цугэ, Т., Ока, А., и Аояма, Т. (2010). Участие Arabidopsis thaliana фосфолипазы Dzeta2 в корневом гидротропизме посредством подавления корневого гравитропизма. Planta 231, 491–497. DOI: 10.1007 / s00425-009-1052-x
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Тонг, Х., Лизер, К. Д., Хоу, Х., Юэн, Г., Бриггс, В., и Хе, З.-Х. (2008). Роль УФ-В зондирования корней в раннем развитии проростков Arabidopsis . Proc. Natl. Акад. Sci. США 105, 21039–21044. DOI: 10.1073 / pnas.0809942106
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Тревавас А. (2014). Поведение и интеллект растений . Оксфорд: Издательство Оксфордского университета. DOI: 10.1093 / acprof: oso / 9780199539543.001.0001
CrossRef Полный текст | Google Scholar
Тревавас А. и Балушка Ф. (2011). Вездесущность сознания. Вездесущность сознания, познания и интеллекта в жизни. EMBO Rep. 12, 1221–1225. DOI: 10.1038 / embor.2011.218
CrossRef Полный текст | Google Scholar
Tsuchiya, Y., Vidaurre, D., Toh, S., Hanada, A., Nambara, E., Kamiya, Y., et al. (2010). Скрининг малых молекул определяет новые функции растительного гормона стриголактона. Nat. Chem. Биол. 6, 741–749. DOI: 10.1038 / nchembio.435
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Умехара, М., Ханада, А., Yoshida, S., Akiyama, K., Arite, T., Takeda-Kamiya, N., et al. (2008). Подавление ветвления побегов новыми терпеноидными гормонами растений. Природа 455, 195–200. DOI: 10.1038 / nature07272
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Уоллер Ф. и Ник П. (1997). Ответ актиновых микрофиламентов во время контролируемого фитохромом роста проростков кукурузы. Protoplasma 200, 154–162. DOI: 10.1007 / BF01283291
CrossRef Полный текст | Google Scholar
Ван, Ю., Jasik, J., Wang, L., Hao, H., Volkmann, D., Menzel, D., et al. (2012). Преобразователь сигнала NPh4 объединяет фотосенсор фототропина-1 с полярным транспортом ауксина на основе PIN2 в корневом фототропизме Arabidopsis . Растительная клетка 24, 551–565. DOI: 10.1105 / tpc.111.094284
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Уотерс, М. Т., Нельсон, Д. К., Скаффиди, А., Флематти, Г. Р., Сан, Ю. К., Диксон, К. В. и др. (2012). Специализация в семействе белков DWARF14 дает различные ответы на каррикины и стриголактоны у арабидопсиса . Разработка 139, 1285–1295. DOI: 10.1242 / dev.074567
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Уотерс, М. Т., Ван, П., Коркарич, М., Кэппер, Р. Г., Сондерс, Н. Дж., И Лэнгдейл, Дж. А. (2009). Факторы транскрипции GLK координируют экспрессию фотосинтетического аппарата Arabidopsis . Растительная клетка 21, 1109–1128. DOI: 10.1105 / tpc.108.065250
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Вернер, Т., Nehnevajova, E., Köllmer, I., Novák, O., Strnad, M., Krämer, U., et al. (2010). Специфичное для корней снижение цитокинина вызывает усиление роста корней, засухоустойчивость и обогащение листьев минералами у Arabidopsis и табака. Растительная клетка 22, 3905–3920. DOI: 10.1105 / tpc.109.072694
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Xu, W., Ding, G., Yokawa, K., Baluška, F., Li, Q.F., Liu, Y., et al. (2013). Усовершенствованный метод на чашках с агаром для изучения роста корней и реакции Arabidopsis thaliana . Sci. Отчет 3: 1273. DOI: 10.1038 / srep01273
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Язданбахш Н., Сюльпис Р., Граф А., Ститт М. и Фисан Дж. (2011). Циркадный контроль удлинения корня и разделения С у Arabidopsis thaliana . Среда растительных клеток. 34, 877–894. DOI: 10.1111 / j.1365-3040.2011.02286.x
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Йокава, К., Кагениши, Т., Кавано, Т., Манкузо, С., и Балушка, Ф. (2011). Освещение корней Arabidopsis вызывает немедленный всплеск продукции АФК. Завод Сигнал. Behav. 6, 1460–1464. DOI: 10.4161 / psb.6.10.18165
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Йонеяма К., Йонеяма К., Такеучи Ю. и Секимото Х. (2007). Дефицит фосфора в красном клевере способствует экссудации оробанхола, сигнала для микоризных симбионтов и стимулятора прорастания корневых паразитов. Planta 225, 1031–1038. DOI: 10.1007 / s00425-006-0410-1
Pubmed Аннотация | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
.